|
|
Post by Admin on Nov 29, 2023 21:45:01 GMT
Next, we decided to further investigate the impact of the amino acid substitutions on the receptor structure. For that, we constructed a three-dimensional (3D) model for MC1R based on the human melanocortin 4 protein receptor (MC4R), with higher sequence identity (>50%) and the muscarinic acetylcholine receptor M2, with higher sequence alignment coverage (0.89). The four structure-based predictions (Table 3, right) are centered on changes in ΔΔG energies that result in positive or negative values for stabilizing or destabilizing effects on the receptor structure, respectively. Ile155Thr, Cys35Tyr and Pro256Ser showed agreeable results regarding destabilization of protein structure with the sequence-based predictors associated with damaging effects (Table 3, left), suggesting that these three mutations would induce negative effects on MC1R by increasing protein flexibility on the mutation sites. The Cys35Tyr mutation could impair the binding of MC1R agonists The nonsynonymous substitution Cys35Tyr was identified in two unrelated red-hair individuals. From the five substitutions selected to be investigated, C35Y presented previous experimental data that indicated a potential role of disrupting a proper function of MC1, directly related to the structure modification. Treatment with reducing agents and induced point mutations on cysteines 35, 267, 273 and 275 have proven that these four cysteine residues are critical [53]. Visual inspection of the 3D structure of MC1R model (Fig 4A) confirmed the two disulfide bond links: Cys267-Cys273 within the extracellular loop 3 (already present on the template MC4R, then transferred to the model) and Cys35-Cys275 between the N-ter and the extracellular loop 3, the latter a potential pair in case the N-terminal loop is in a closer position.  Fig 4 Role of the Cys35-Cys275 disulfide bridge on the MC1R structure dynamics. (A) MC1R structure model is shown as cartoon colored in cyan and four critical cysteine residues are represented as sticks, colored in pink. The extracellular loop 3 is highlighted in dark blue. The disulfide bridge formed by residues 267 and 273 was derived from the MC4R structure used as template for comparative modelling of MC1R. Cys35 and Cys275 could be a potential pair for a second disulfide bridge on the protein structure. (B) Root Mean Square fluctuation (RMSF) for MC1R residues in presence of the disulfide bond (Cys35-Cys275), in its absence (Cys35) and upon Cys35Tyr mutation. (C) MC1R binding site volume variation along the MD simulation trajectories. The plot accounts for both MD replicas of MC1R model in presence of the disulfide bridge (bridge) and in absence of the disulfide bridge (control). Median values are indicated on the plot. (D) MC1R conformational changes observed during MD simulations. Representative structures for each system are displayed as a cartoon, colored in cyan. The N-ter first 15 residues are colored in red and the mutation site is indicated as a red sphere. The binding pocket is represented as surface, colored in light orange. When MC1R model bears the Cys35-Cys275 disulfide bond (top), the accessibility of the binding pocket is maintained, with no interference of the N-terminal loop; while in the absence of the bond (middle), the N-terminal loop of MC1R is maintained at the edge of the binding cavity area, interfering with its accessibility. Finally, upon mutation to tyrosine (bottom), the structure suffers an important conformational change that decreases dramatically the pocket volume, and therefore, its capacity to welcome binders. Considering the importance of these post-translational modifications in MC1R function, we investigated MC1R under three different scenarios: Cys35-Cys275: disulfide bond present; Cys35: disulfide bond absent; Cys35Tyr: disulfide bond absent and mutation on position 35. Molecular dynamics (MD) simulations were performed for the three MC1R model structures. Root Mean Square Fluctuations (RMSF) is a measure of residue flexibility over the course of the MD simulations. We notice that the absence of the disulfide bond Cys35-Cys275 or the mutation Cys35Tyr increases the overall flexibility of the protein when compared with the structure with the disulfide bond (Fig 4B), making the Cys35Tyr mutant significantly more flexible than the other two systems. This shows that the disulfide bond is important to stabilize the overall protein conformation and the mutation to tyrosine is impacting the overall structure stability. To further investigate the impact of this increased flexibility on MC1R function, we estimated the average volume of the binding pocket to see if the mutation could impact the binding site region. On the Cys35-Cys275 model, the cavity presents a slightly bigger volume when compared with Cys35, where the disulfide bond is absent (Fig 4C). However, upon mutation to tyrosine, the MC1R binding pocket has a dramatic reduction on the average volume, confirming that the mutation is indeed provoking a conformational change on the receptor that has a significant impact on the integrity of the binding site. Visual inspection of the MD trajectories and representative structures show that the pocket accessibility is reduced in the absence of the disulfide bond due to the presence of the N-ter loop consistently at the edge of the binding pocket cavity (Fig 4D). Upon mutation to tyrosine, the N-ter loop is not interfering with the binding pocket accessibility, however the increased structure flexibility deforms the overall fold of the receptor and consequently diminishes the binding pocket volume (Fig 4D). This indicates that Cys35-Cys275 is not only important for the structural integrity of the receptor but also to its function by maintaining the binding pocket accessibility and the Cys35Tyr mutation disrupts the functional conformation of MC1R receptor. In the absence of the disulfide bond, there is an increase of the N-terminal loop hydrogen bonds interactions with the binding pocket edge residues and adjacent loops, while upon mutation there is an increase of the overall receptor flexibility that compromise the receptor fold (S1 Movie). These data suggest that the negative effect associated with the SNP Cys35Tyr might be the rearrangement of the receptor structure disfavoring the access to MC1R binders. |
|
|
|
Post by Admin on Nov 30, 2023 21:36:44 GMT
Discussion
This work characterized the entire coding region of melanocortin 1 (MC1R) receptor gene in an admixed population from Southeast of Brazil. The MC1R is widely studied in humans and non-humans and several variants have been described to influence melanin production [16]. We detected 31 different polymorphisms comprehending synonymous, nonsynonymous substitutions and indels, which agrees with the notion that the MC1R is considered a polymorphic gene [54]. The variants associated with damaging effects are commonly genetic alterations that impair the protein function by changing the codon reading frame (indel mutations) or by a single nucleotide substitution resulting in a different codon that produces an amino acid change (nonsynonymous). Based on the knowledge that MC1R is better related to red-hair/fair skin phenotype within Europeans or European descendants, in our population all the red hair individuals carry at least one polymorphism known or predicted as loss-of-function variant and, with the exception to Arg151Cys observed in one individual, all the alleles are in heterozygous state. Considering the phase of the alleles is unknown, we cannot affirm whether the MC1R mutation in these groups would be sufficient to explain the phenotype.
As mentioned, the MC1R gene is highly polymorphic and several altered alleles exhibited low frequency, making any robust correlation analysis between genotype and phenotype difficult. Among the several rare alleles detected in our sample population, three were novel synonymous substitutions: Leu11Leu, Tyr143Tyr and Ala181Ala. Probably, these three substitutions appeared in Brazilian population since the genetic diversity of MC1R has been extensively investigated and none of them has been identified in any previous studies or in parental origin populations.
Several theories have been proposed to explain the differences in polymorphic contents on MC1R between European and African populations, and the most acceptable is related to ultraviolet radiation b (UVb) exposure [4]. According to Jablonski team, the UV incidence is inversely correlated to latitude; in a constraint pressure towards the maintenance of dark skin in low latitude whereas directional selection is observed in high latitude regions making the skin lightening more advantageous throughout human evolution. A health balance should be achieved in a way to find fine control mechanisms in order to protect the folate from photolysis while at the same time allowing the synthesis of vitamin D. The NMDS based on the genetic diversity of MC1R comprising 942bp in length (GRCh37 16:89985667–89986608) was able to efficiently discriminate the three main regions: Africa, Asia and Europe (Fig 2). Although Asians have low levels of melanin on the skin compared to Africans, the lightening of skin occurred as an independent evolutionary process as already demonstrated by other groups [55, 56]. None of the three Brazilian skin color phenotypes clustered to Asians, and this has been supported by the low frequency of Asian matrilineal ancestry (6.7%) in our population. Considering that the significant immigration from Asia to Brazil occurred in the 19th century, it is a recent event in terms of genetic admixture.
As mentioned above, according to the skin reflectance, consequently the Africans are characterized as having dark skin, whereas Europeans are the light ones. The ordination method showed that dark skin group were clustered among Africans explained by the intense traffic of black Africans during the Atlantic slave trade period, especially those from the Bantu ethnic group [57, 58]. Regarding the light skin color, this group gathered with Europeans, in particular Mediterranean populations, in line with the colonization of Rio de Janeiro [20]. Despite the gene flow among Native Americans, Africans and Europeans, the Fst value of light and intermediate skin color was very close to that of European populations, indicating similar genetic background in terms of skin color based on MC1R (Fig 2 and S3 Table). These findings are not related with mtDNA analysis, in which higher African ancestry was observed in 53% of individuals of intermediate skin, therefore not reflecting the distribution of skin color (Fig 3).
It has already been described that the matrilineal ancestry has an equivalent proportion of European, African and Native American haplogroups among Brazilians, more precisely in Southeastern region [59]. This diversified ancestry profile is present when the total sample population of RJ was analyzed (Fig 3), validating the representativeness of our population of study. However, when the RJ population was categorized based on skin color phenotype, the mtDNA haplogroup distribution was in agreement with the corresponding European and African ancestry origin for light and dark populations, respectively. This is in agreement with the notion that skin color variation in the new world colonies is a product of parental population contribution [12] however, the same conclusion was not reached regarding the intermediate skin color phenotype. Despite the intermediate group being clustered together with light skin populations, the European haplogroup represented only 20% with major contribution from Africans (53%), indicating that the color variation could not be totally explained by matrilineal ancestry inheritance. Analysis using the ancestry-informative markers (AIMs) from somatic chromosomes would be a better strategy for understanding the complexity of color variation in our admixed population.
Based on this AIMs approach, a previous study evaluated several aspects of physical traits in admixed populations from Latin American [23]. Individuals categorized as mixed skin color showed a high percentage of European genetic ancestry (0.69), probably due to the mean melanin index (MI) being more prone to the white spectrum than the black one. It is comprehensible due to the southern region of Brazil having historically received many European immigrants [19, 22] compared to Rio de Janeiro (Southeast), which had higher contribution of African and Native American as demonstrated by another group [20]. However, even considering the colonization particularities between both regions, the intermediate phenotype remains closer to light group as previously observed in GWAS study [23].
Several nonsynonymous polymorphisms present on the coding region of MC1R have already been subjected by in silico predictors or by in vitro assays to evaluate their impacts on receptor function [13, 14]. In this work we have chosen five SNPs; three found in blond-red hair individuals (Cys35Tyr, Ile155Thr and Pro256Ser) and two in black skin individuals (Val156Leu and Phe196Leu). According to the ClinVar database, the SNPs Ile155Thr, Val156Leu and Phe196Leu have been already described as natural variants. ClinVar reports Ile115Thr (VCV000239154.7) as having conflicting interpretations of pathogenicity, Val156Leu (VCV000258651.4) and Phe196Leu (VCV000321433.4) as benign/likely benign, whereas Cys35Tyr (VCV000430181.6) and P256S (VCV000470711.4) showed uncertain significance. Based on these discordant outcomes, we decided to estimate the possible phenotypic effects for all of them using seven disease-association predictors based on the amino acid sequence of the protein. The variants identified in fair skin individuals were predicted to promote negative effect on function as well as destabilize MC1R structure (Table 3). The destabilizing phenotypes are supported by the nature of amino acid substitutions on protein structures. Considering the isoleucine replacement for a threonine could increase polarity and contribute to structure rigidity since its side chain movement is more limited, this could be the reason for the observed phenotype. A meta-analysis on MC1R gene showed a significant association between Ile155Thr and the development of melanoma that were corroborated by the fact that this mutation impaired cAMP signaling [60, 61]. Also, Ile155Thr was individually associated with malignant melanoma risk in the Spanish population [62]. Larger, prospective cohorts with different ethnic backgrounds are needed to verify the MC1R variant associations. Regarding Pro256Ser, the proline replacement for a serine on the core of the alpha helix could increase flexibility on this region. Additionally, proline residues in the transmembrane (TM) alpha-helices of integral membrane proteins have long been suspected to play a key role for helix packing and signal transduction by inducing regions of helix distortion and/or dynamic flexibility (hinges), strengthening the idea of Pro256Ser exhibiting a loss-of-function effect. Proteins vary in stability and a ΔΔG in the range of 2 kcal/mol is considered to result in a mutational “hot spot” of sufficient effect [63] on its conformation, required for protein’s function in vivo. The predicted changes on protein stability for both mutations have energy values that are inferior or around –2.0 kcal/mol (Table 3), so further studies need to be conducted.
Cys35Tyr was predicted by sequence and structure-based methods as a damaging/destabilizing mutation (Table 3). In addition, previous experimental data have proven that four cysteine residues are critical for the proper function of MC1R, since the treatment with reducing agent DTT as well as induced point mutations on cysteines 35, 267, 273 and 275 impair both ligands interaction with MC1R and cAMP signaling, indicating that disulfide bridges are required for the integrity of the receptor [53].
The role of disulfide bonds has been investigated for the rhodopsin-like family. One disulfide bridge between the extracellular loop 2 and the transmembrane helix 3 is very conserved and increases the constraint on the protein structure [64]. Other disulfide bridges have also been observed, concerning the N-terminal and the extracellular loop 3 for instance on the CXCR4 receptor [65] and within the extracellular loop 3 for human A2A adenosine receptor, melanocortin receptors and human histamine H1 receptor [64], as could be the case for the Cys35 (N-terminal) and Cys275 (extracellular loop 3) on MC1R.
MD simulations of MC1R in absence of the Cys35-Cys275 disulfide bridge have shown that the N-terminal can be flexible and interact with other protein residues, impairing the access to the binding pocket cavity (S1 Movie). Interestingly, MC4R has a tyrosine residue on the same position in sequence for Cys35 at MC1R and the N-terminal portion of the MC1R structure could not be modelled using the MC4R crystal structure as a template, since the equivalent region was missing, which could indicate that it is a very flexible region. We could also assume that an increase of flexibility on this region is expected on MC1R upon mutation.
Our MD simulations of the mutated Cys35Tyr receptor confirmed that hypothesis: tyrosine induces an increase on the overall flexibility of the receptor, leading to a conformational change that dramatically reduces the binding pocket volume. This conformational change could potentially impair the binding of agonists and therefore MC1R function (Fig 4).
Two possibilities could be drawn: a conformational change responsible by impairing agonist binding or the new residue would promote a different signaling, for instance, an antagonist-like signal. The latter hypothesis corroborates previous data on the loss-of-function effect observed in vitro, the retention of MC1R in the endoplasmic reticulum [53, 66]. Although the change from cysteine to tyrosine being a natural substitution instead of alanine [66] obtained by mutagenesis, we are not able to confirm whether Cys35Tyr could produce the same outcome, since tyrosine is well known for being phosphorylated and another signaling could be triggered.
In conclusion, we evaluated the genetic diversity of melanocortin-1 receptor (MC1R) in an admixed population from Rio de Janeiro. Analysis based on MC1R variants distribution grouped RJ population light and intermediate skin color with Europeans, despite the intermediate matrilineal ancestry being comprised of majority African origin. The gene exhibited a high polymorphic profile among the light skin group with prominent proportion of nonsynonymous substitutions. Among them, several loss-of-function mutations were detected in red-hair individuals. We demonstrated through MD simulations that the Cys35Tyr mutation potentially impairs agonist binding due to the disruption of the disulfide bond Cys35-Cys275 and induced conformational changes on the receptor structure impacting its binding site. This work has shed light on the mutations associated with the MC1R gene and contributed to a better understanding of the genetic diversity of MC1R in an admixed population with a wide range of color variants.
|
|
|
|
Post by Admin on Dec 12, 2023 3:09:50 GMT
Genomic history of coastal societies from eastern South America Abstract Sambaqui (shellmound) societies are among the most intriguing archaeological phenomena in pre-colonial South America, extending from approximately 8,000 to 1,000 years before present (yr BP) across 3,000 km on the Atlantic coast. However, little is known about their connection to early Holocene hunter-gatherers, how this may have contributed to different historical pathways and the processes through which late Holocene ceramists came to rule the coast shortly before European contact. To contribute to our understanding of the population history of indigenous societies on the eastern coast of South America, we produced genome-wide data from 34 ancient individuals as early as 10,000 yr BP from four different regions in Brazil. Early Holocene hunter-gatherers were found to lack shared genetic drift among themselves and with later populations from eastern South America, suggesting that they derived from a common radiation and did not contribute substantially to later coastal groups. Our analyses show genetic heterogeneity among contemporaneous Sambaqui groups from the southeastern and southern Brazilian coast, contrary to the similarity expressed in the archaeological record. The complex history of intercultural contact between inland horticulturists and coastal populations becomes genetically evident during the final horizon of Sambaqui societies, from around 2,200 yr BP, corroborating evidence of cultural change. Main The settlement of the Atlantic coast by maritime societies is a central topic in South American archaeology. Across ~3,000 km of the coast of Brazil, semi-sedentary populations, with seemingly large demography, produced thousands of shellmounds and shell middens, locally known as sambaquis (heaps of shell, in the Tupi language), for over 7,000 years. Subsistence was based on a mixed economy, combining aquatic resources and plants, complemented by hunting of terrestrial mammals and horticulture1,2,3,4,5,6,7,8. Sambaquis are the product of planned and long-term deposition of shells, fish remains, plants, artefacts, combustion debris and local sediments, and they were used as territorial markers, dwellings, cemeteries and/or ceremonial sites. On the southern Brazilian coast, funerary shellmounds can reach monumental heights (of up to 30 metres) and often contain hundreds of human burials, suggesting a high demographic density unparalleled in the South American lowlands3,6,9,10,11. In a singular enclave south of São Paulo State, further inland from the coast (Vale do Ribeira de Iguape), sambaqui sites are within the Atlantic Forest12,13,14,15. Here there is evidence of early Holocene settlement in the riverine sambaqui of Capelinha, as revealed by a male individual directly dated to ~10,400 years before present (yr BP) (we identify all analysed individuals by rounding the mean calibrated age in years BP)10. This individual was named ‘Luzio’, as a reference to ‘Luzia’, a final Pleistocene female skeleton found in the Lagoa  region in east-central Brazil10,16,17. Both individuals are at the centre of long-lasting debates for exhibiting the so-called paleoamerican cranial morphology that differs from that of present-day indigenous peoples10,18. The earliest evidence of human settlement on the Atlantic coast starts between ~8,700 and 7,000 yr BP with an intensification of sambaqui construction between 5,500 yr BP and 2,200 yr BP2,4,6,19. The relationship between riverine and coastal sambaquis is still a matter of debate, although bioarchaeological studies point towards a biological link20,21,22,23, and some researchers suggest a late Pleistocene/early Holocene cultural connection that faded through time24,25,26,27. The disappearance of Sambaqui societies started 2,000 years ago, when funerary fishmounds replaced shellmounds in the territory where they previously thrived4,28,29,30,31. This abrupt change in the archaeological record is concomitant with environmental and ecological changes related to coastal regression and climatic events32,33,34,35,36 that had an irreversible impact on the availability of key resources. Between 1,200 and 900 years ago, thin-walled non-decorated pottery (Taquara-Itararé tradition) appeared for the first time on the southern Brazilian coast2,6,9,11,29,37,38,39. The makers of Taquara-Itararé ceramics were horticulturists that arrived in the southern Brazilian highlands about 3,000 years ago, lived in pit houses and cremated their dead in funerary mounds. They are considered to be the ancestors of present-day Jê-speaking indigenous peoples of southern Brazil (Kaingang, Xonkleng, Laklãnõ and the extinct Kimdá and Ingáin), a language family of the Macro-Jê stock38,40,41,42,43. The dispersal of Taquara-Itarare ceramics on the southern coast was first interpreted as resulting from the demographic expansion of inland horticulturists. However, evidence points to a complex scenario of social interaction between inland and coastal populations, with changes in funerary practices and post-marital residence patterns after the introduction of ceramics, biological continuity and maintenance of mobility patterns (with local variations), persistence in the exploitation of aquatic resources, and development of sophisticated fishing technologies2,4,11,21,23,39,44,45,46,47,48,49. Ceramics appear in the southeast coast about 2,000 years ago but are associated with the Una tradition, also probably produced by speakers of the Macro-Jê language stock50,51. Shortly after the appearance of southern proto-Jê ceramics, another major transformation occurred on the Atlantic coast. This is documented by the arrival of speakers of the Tupi-Guarani language family (of the Tupi stock), a forest-farming culture who migrated from southern Amazonia more than 2,500 years ago in one of the largest expansion events in the indigenous history of South America. Although still a matter of debate, the Tupi-Guarani would have dispersed southwards from southwestern Amazonia (homeland of the Tupi stock) across the core of South America, reaching the La Plata basin, and almost simultaneously from southeastern Amazonia across the Atlantic coast of Brazil38,42,52,53,54,55,56,57. While on the southern coast of Brazil a late Tupi-Guarani chronology is well defined38,52, on the southeast coast a much earlier arrival (~3,000 years ago) has been proposed on the basis of the archaeological record of the Araruama region (Rio de Janeiro State)58,59,60. European colonists encountered thousands of Tupi-Guarani peoples both on the Atlantic coast and along major rivers and their tributaries in southern Brazil and northeastern Argentina (Paraná, Paraguay and Uruguay river basins). The Tupi-Guarani produced painted ceramics (red and black on white painting), applied a diversity of plastic decorations and made pots with complex and composite contours that are archaeologically defined as Tupiguarani, Tupinambá and Guarani, depending on the geographical location42,53,61. Ancient DNA data from Brazil are still very sparse, with only 19 published individuals with analysable genomic coverage62,63. Early Holocene individuals from Lapa do Santo in the Lagoa region, dated between ~9,800 and 9,200 yr BP, carried a distinct affinity to the oldest North American genome, which is associated with the Clovis cultural complex (Anzick-1, ~12,800 yr BP)63,64. A genetic signal of 3–5% Australasian ancestry—known as the Population Y signal—was found in present-day indigenous individuals from southwestern Amazonia, Central Brazil and the northwestern South American coast65,66 and in one early Holocene individual from Lapa do Sumidouro (Sumidouro 5, dated to c. 10,400 yr BP)62. However, this signal was not detected in the early Holocene burials from Lapa do Santo, located only four kilometres from Lapa do Sumidouro63. The complete absence of ancient DNA data for Amazonia and Northeast Brazil and the low-coverage data from the south/southeast Brazilian coast have prevented an assessment of whether the Population Y signal survived in those regions through time. Regarding Sambaqui societies, three previously published middle Holocene individuals from Laranjal and Moraes (both riverine shellmounds from the southeast coast of Brazil) and five individuals from the late Holocene site of Jabuticabeira II (one of the largest coastal shellmounds in southern Brazil) showed some level of genetic continuity with present-day indigenous populations63. The analysed Jabuticabeira II individuals carried a significant affinity to present-day Kaingang (Jê speaking) from the southern Brazilian highlands. Although based on low-coverage genome-wide data, this supports a shared ancestry between the Sambaqui societies and the speakers of proto-Jê63. The long-term permanence, cultural similarity and rapid disappearance of Sambaqui societies, plus their archaeological and seemingly genetic disconnection from early Holocene hunter-gatherers, raise numerous questions about their origins and demographic history. First, were Sambaqui individuals genetically different from hunter-gatherers from the hinterland (for example, east-central and northeastern Brazil)? Second, were the riverine Sambaqui groups genetically related to the ones on coastal sites? Third, was there genetic homogeneity across Sambaqui groups from the south and southeast coast of Brazil? Fourth, was the demise of sambaqui construction after 2,000 yr BP and the appearance of ceramics associated with an intensification of contacts with inland populations? Finally, are there genetic connections between Sambaqui groups and other archaeological and present-day indigenous populations from Amazonia and central and northeastern Brazil? www.nature.com/articles/s41559-023-02114-9. |
|
|
|
Post by Admin on Dec 13, 2023 3:18:27 GMT
Results Dataset and ancient DNA authenticity To understand the genetic structure of pre-colonial Brazilian groups and assess their potential genetic transformations through time, we attempted to retrieve ancient DNA from 82 individuals from 24 archaeological sites across four regions: the southeastern and southern Atlantic coast, Lagoa , the lower Amazon, and northeastern Brazil (Supplementary Information and Supplementary Data 1). After applying established criteria for ancient DNA authentication, we obtained a final dataset of genome-wide data from 34 individuals from 11 archaeological sites spanning the past ~10,000 years (Fig. 1 and Supplementary Data 1). We produced genome-wide data via in-solution capture by enriching for a targeted set of ~1.24 million single nucleotide polymorphisms (SNPs) across the human genome (1240k SNP capture)67. We also captured the entire mitochondrial genome (mtDNA) to assign mtDNA haplogroups and to estimate contamination levels, which were found to be low for all cases (<2%). Nuclear DNA contamination estimated for 20 male individuals on the basis of X-chromosome heterozygosity levels68 was also low (<3.5%). Principal component analysis (PCA) and a cluster analysis including worldwide populations further confirmed that all individuals fall within Native American genetic diversity (Extended Data Figs. 1 and 2). For population genetic analyses, we combined the newly authenticated ancient Brazilian genome-wide dataset with previously published ones62,63. Individuals were grouped on the basis of archaeological site, radiocarbon date and genetic affinities established through f3 outgroup statistics (Methods and Supplementary Data 1). Fig. 1: Geographic and temporal distribution of analysed genome-wide data from Brazil. 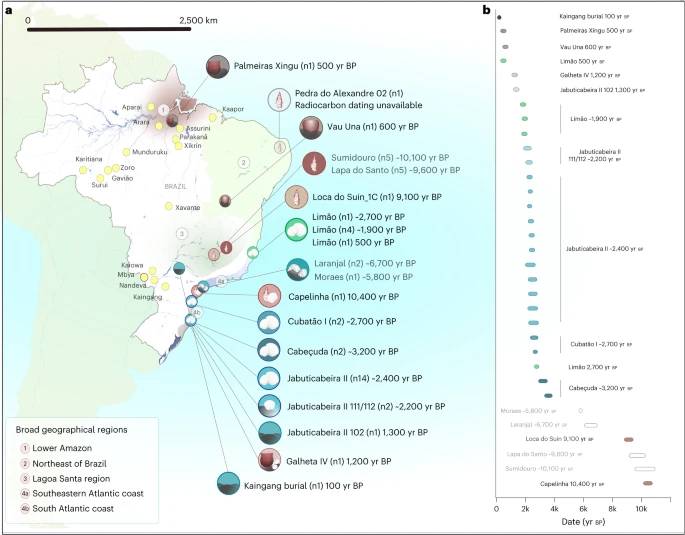 a, The archaeological sites analysed in this study, with the number of analysed individuals reported in brackets. Sites with newly reported genome-wide data are shown in black font, and those with previously published genome-wide data are shown in grey (this color scheme is maintained in all main text figures). The symbols used for each site refer to the associated archaeological cultures (see the legend in Extended Data Fig. 8). The shaded areas represent the broad geographic regions analysed in this work: (1) lower Amazon, (2) northeastern Brazil, (3) Lagoa , (4a) southeastern Atlantic coast and (4b) southern Atlantic coast. The Kaingang burial is geographically closer to the southeastern Atlantic coast but was included in the southern Atlantic group due to its specific genetic affinity. The locations of present-day indigenous groups are represented with yellow dots. b, The calibrated ages (coloured bars) of single directly dated individuals with new genomic data and, in black font, the mean calibrated ages for the respective groups/individuals. For the previously published ancient genome-wide data62,63, the mean calibrated ages for the respective groups/individuals are reported in grey, whereas the white bars represent the temporal range of all directly dated individuals included in each group. Figure related to Supplementary Data 1. |
|
|
|
Post by Admin on Dec 13, 2023 21:53:53 GMT
Early Holocene hunter-gatherer radiation The oldest human presence in southeastern Brazil is directly attested by the ‘Luzio’ individual, a skeleton buried in the riverine shellmound of Capelinha genetically analysed here (Capelinha_10400BP). The morphological similarity of this male individual to paleoamerican features observed in early Holocene groups from the Lagoa region, and the chronological gap of almost 3,000 years with other burials from the same site, call into question his association with riverine Sambaqui societies10. We investigated the genetic affinities of Capelinha_10400BP to other ancient Brazilian individuals using f4 statistics of the form f4(Mbuti, Capelinha_10400BP; ancient Brazilians—left, ancient Brazilians—right) (Extended Data Fig. 3a and Supplementary Data 2). None of the tested ancient individuals show a higher allele sharing with Capelinha_10400BP, even when the temporally close and phenotypically similar Lagoa groups are considered. The same pattern is observed when Capelinha_10400BP is compared with an early Holocene hunter-gatherer from Loca do Suin, dated to ~9,100 yr BP and located 200 km southwest of the Lagoa region (Loca do Suin_9100BP). Conversely, the Lapa do Santo_9600BP and Sumidouro_10100BP groups share a higher genetic affinity with each other than with any other ancient Brazilian group (significance considered at Z > |3|, if not differently indicated) (Extended Data Fig. 3a). These results indicate that Capelinha_10400BP does not represent an early occupation of the southeast coast by inland groups carrying Lagoa –related ancestry and suggest that his population did not leave a substantial genetic contribution in the later Brazilian individuals analysed here. We then used qpWave69 to estimate the minimum number of streams of ancestry necessary to explain the genetic variation observed among early Holocene hunter-gatherers across South America. Our results show that Capelinha_10400BP and Loca do Suin_9100BP cannot be distinguished from other early Holocene populations as part of a distinct wave of ancestry (P > 0.01) (Supplementary Data 3). To the limit of our resolution, the lack of close affinity among early Holocene individuals from different South American sites suggests that they derived from a rapid radiation event63. A previous study also revealed that the oldest South American genomes, Los Rieles_11900BP from Chile and Lapa do Santo_9600BP from Brazil, carried a higher affinity to the Clovis-associated Anzick-1 individual from North America than Lauricocha_8600BP from Peru did63. With f4 statistics, we could show that while Capelinha_10400BP and Sumidouro_10100BP do not have a lower affinity to Anzick-1 than Los Rieles_11900BP and Lapa do Santo_9600BP do, they also do not show a higher affinity to Anzick-1 than Lauricocha_8600BP does (Supplementary Data 4). To measure the relative proportion of the Anzick-1-related ancestry in ancient South American groups, we performed an f4-ratio test70 (Methods), using Los Rieles_11900BP and Lauricocha_8600BP as the reference individuals with the maximum and minimum amount of such ancestry in early Holocene South America, respectively. Our results corroborate that Lapa do Santo_9600BP carry a significantly higher amount of Anzick-1-related ancestry than Lauricocha_8600BP (Z = 3.31), while the other tested groups show different proportions without reaching significance (Extended Data Fig. 4 and Supplementary Data 4). This trend suggests a genetic gradient of Anzick-1-related contribution in early South American hunter-gatherers rather than a scenario of two isolated migration waves with and without Anzick-1-related ancestry. Shellmound societies from the middle to the late Holocene To investigate the affinities between riverine and coastal Sambaqui groups, we analysed our newly produced data alongside previously published individuals from the riverine sambaquis Laranjal (n = 2, ~6,700 yr BP) and Moraes (n = 1, ~5,800 yr BP)63 (Extended Data Fig. 3b). The southeast coast shellmounds are represented by the sambaqui do Limão (n = 6, ~2,700–500 yr BP), located in the State of Espírito Santo. The south coast Sambaqui are represented by individuals from three shellmounds—Jabuticabeira II (n = 17, ~2,500–1,300 yr BP), Cabeçuda (n = 2, ~3,200 yr BP) and Cubatão I (n = 2, ~2,700–2,600 yr BP)—and one individual from the fishmound Galheta IV (~1,200 yr BP), representing the final horizon of Sambaqui societies. Our analyses confirm the strong local genetic affinity between the riverine Sambaqui individuals compared with all other ancient Brazilian groups in our dataset (f4(Mbuti, Laranjal_6700BP; ancient Brazilian group, Moraes_5800BP) > 0). Individuals from the riverine sites also show genetic similarities to individuals from the southern coastal sambaquis of Cubatão I (CubatãoI_2700BP), Cabeçuda (Cabeçuda_3200BP) and Jabuticabeira II (JabuticabeiraII_~2400BP), indicating some level of genetic continuity through time between riverine shellmound builders and Sambaqui societies from the southern coast. Interestingly, this genetic similarity is not observed between the riverine shellmounds and the sambaqui do Limão, located further north (Fig. 1 and Extended Data Fig. 3b). To improve our knowledge on the genetic interactions among Sambaqui groups, we co-analysed all individuals from the five coastal sites, which are located up to 1,500 km apart along the southeast and south coasts (Fig. 1). The archaeological site with the largest number of analysed genome-wide data is Jabuticabeira II. The 17 individuals from this site cluster in three genetically distinct groups, as revealed through f3 and f4 tests (Supplementary Data 2): (1) a main cluster, composed of 14 individuals dated to ~2,500–2,300 yr BP (JabuticabeiraII_~2400BP—we identify genetic groups by rounding the mean calibrated age for all dated individuals; Supplementary Data 1), of which 12 are not first degree related and are grouped together for analysis; (2) two first-degree-related individuals dated to ~2,200–2,100 yr BP (JabuticabeiraII_111/112_~2200BP, only one individual used for analysis); and (3) the most recent individual, dated to ~1,300 yr BP (JabuticabeiraII_102_1300BP). This skeleton was found in the topmost shell deposit and exhibited a different funerary pattern from the older burials, including an extended rather than a flexed position and the absence of grave goods (Supplementary Data 1). In f4 statistics, we found a higher genetic affinity between the three groups from Jabuticabeira II compared with all other ancient Brazilian groups (Supplementary Data 2). The temporally intermediate individual appears to be genetically intermediate to the preceding and succeeding individuals, as indicated by f4(Mbuti, JabuticabeiraII_111/112_~2200BP; JabuticabeiraII_~2400, JabuticabeiraII_1300BP) ~ 0 (Z = 0.47). Intersite comparisons showed higher allele sharing between the JabuticabeiraII_~2400BP group, JabuticabeiraII_111/112_~2200BP, Cabeçuda_3200BP and GalhetaIV_1200BP, to the exclusion of other ancient Brazilian groups. The CubatãoI_~2700BP group shows genetic connections to the other southern shellmound groups such as JabuticabeiraII_~2400BP and Cabeçuda_3200BP. The affinities between these four shellmound and fishmound individuals thus reveal the presence of a late Holocene genetic cluster in the southern coast of Brazil (Fig. 2). |
|


 region in east-central Brazil10,16,17. Both individuals are at the centre of long-lasting debates for exhibiting the so-called paleoamerican cranial morphology that differs from that of present-day indigenous peoples10,18. The earliest evidence of human settlement on the Atlantic coast starts between ~8,700 and 7,000 yr BP with an intensification of sambaqui construction between 5,500 yr BP and 2,200 yr BP2,4,6,19. The relationship between riverine and coastal sambaquis is still a matter of debate, although bioarchaeological studies point towards a biological link20,21,22,23, and some researchers suggest a late Pleistocene/early Holocene cultural connection that faded through time24,25,26,27.
region in east-central Brazil10,16,17. Both individuals are at the centre of long-lasting debates for exhibiting the so-called paleoamerican cranial morphology that differs from that of present-day indigenous peoples10,18. The earliest evidence of human settlement on the Atlantic coast starts between ~8,700 and 7,000 yr BP with an intensification of sambaqui construction between 5,500 yr BP and 2,200 yr BP2,4,6,19. The relationship between riverine and coastal sambaquis is still a matter of debate, although bioarchaeological studies point towards a biological link20,21,22,23, and some researchers suggest a late Pleistocene/early Holocene cultural connection that faded through time24,25,26,27.