|
|
Post by Admin on Dec 14, 2023 20:03:11 GMT
Fig. 2: f4 statistics among ancient Brazilian groups and/or individuals from the southern and southeastern coasts dated between 3,000 yr BP and 1,000 yr BP. 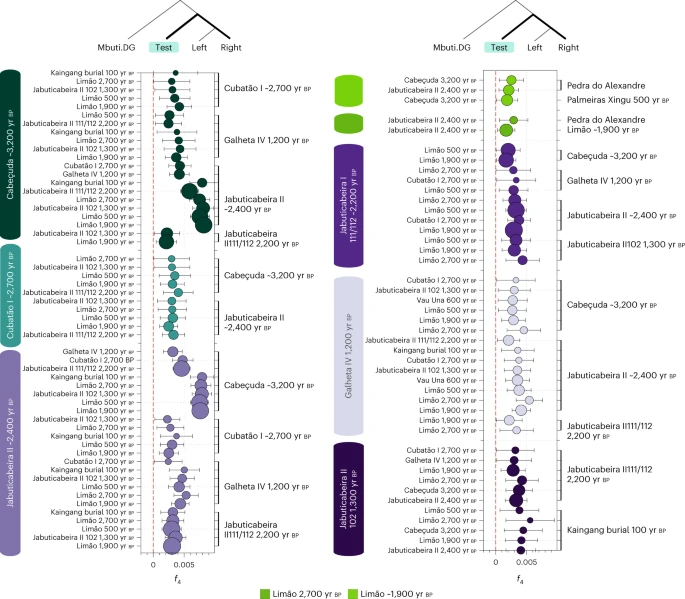 Significant f4 statistics (Z > 3) polarized to positive values performed on ancient Brazilian genome-wide data in the form f4(Mbuti, TEST; ancient Brazilians—left, ancient Brazilians—right) to test allele sharing among the different groups using the 1240k dataset. The point sizes refer to the number of SNPs used to compute the f4 tests (at least 20,000 SNPs), and the colours correspond to the TEST group/individual. The bars represent f4 statistics ± 3 standard errors. Figure related to Supplementary Data 2. On the southeastern coast, the genetic similarities ascertained through f3 outgroup and f4 statistics revealed three distinct groups at the sambaqui do Limão: (1) the oldest individual (Limão_2700BP), (2) a cluster of four temporally intermediate individuals (Limão_~1900BP) and (3) the most recent individual (Limão_500BP). The Limão_~1900BP group shows the highest genetic affinities to Limão_2700BP and to a possibly early to middle Holocene hunter-gatherer from the northeastern site of Pedra do Alexandre (Pedra Do Alexandre2_undated)71 (Fig. 2). This result demonstrates genetic connections between Sambaqui individuals from the southeast coast and hunter-gatherer groups from northeastern Brazil. When combined with the results obtained from the south coast sambaquis, our analyses indicate that shellmound societies from the south and southeast (that is,  Catarina and Espírito Santo states, respectively) do not constitute a genetically homogenous population, as previously suggested by the analyses of cranial and dental morphological variation21,23. |
|
|
|
Post by Admin on Dec 17, 2023 0:17:46 GMT
The final horizons of shellmound societies The significance of Taquara-Itararé ceramics (associated with proto-Jê speakers) at coastal sites after the final horizon of sambaqui construction has been at the centre of recent academic debates. According to some scholars, an intensification in contacts with proto-Jê-speaking groups after ~2,000 yr BP, even before the appearance of ceramics at the coast, would have led to the demise of Sambaqui societies9,11,30. In this work, the post-2,000 yr BP horizon is represented by JabuticabeiraII_102_1300BP (Fig. 2), buried at the top of the shell deposit, and by an individual from Galheta IV, a fishmound with Taquara-Itararé pottery (GalhetaIV_1200BP) (Fig. 2). To further investigate the genetic connections between individuals in sambaquis and fishmounds, proto-Jê-speaking groups, and present-day indigenous peoples, we merged our ancient genomic data with two published present-day genomic datasets: (1) the Illumina dataset assembled in Reich et al.72, combined with 1240k SNP capture data generated in this study from an early twentieth-century southeastern Kaingang individual from the state of São Paulo (Kaingang burial_100BP), showing distinctive affinity with present-day southern Kaingang; and (2) the Human Origins dataset54,66,69,73. Using the Illumina dataset, we observed patterns of shared genetic drift between some Sambaqui groups and present-day Kaingang (Fig. 3a and Extended Data Fig. 5). To formally test this affinity, we performed the following f4 tests: (1) f4(Mbuti, ancient coastal group; Kaingang, other present-day indigenous groups) and (2) f4(Mbuti, Kaingang; ancient coastal group A, ancient coastal group B). The results from the first test reveal an excess of genetic similarity between present-day Kaingang and JabuticabeiraII_102_1300BP. The second test expands this finding by showing that JabuticaberiaII_111/112_~2200BP and even more so JabuticabeiraII_102_1300BP are genetically closer to present-day and twentieth-century Kaingang, when compared with not only the JabuticabeiraII_~2400BP group but also the Taquara-Itararé-pottery-associated GalhetaIV_1200BP individual (Supplementary Data 5 and Extended Data Fig. 5). This genetic link between Kaingang and the younger Jabuticabeira II individuals corroborates the hypothesis of an intensification of contacts between proto-Jê-speaking groups and Sambaqui societies of the southern coast, at least from ~2,200 yr BP. Fig. 3: Heat map of f3 outgroup statistics. 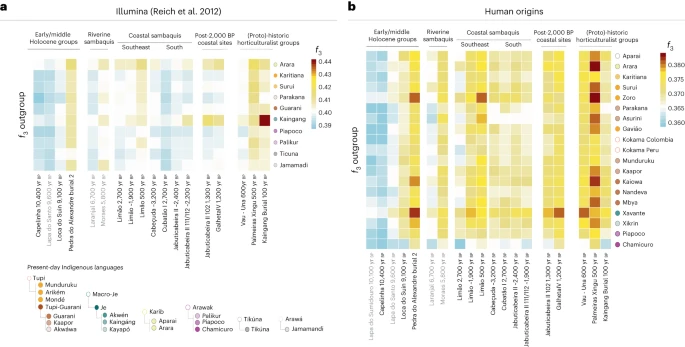 a, Comparisons between ancient and present-day groups/individuals using the Illumina dataset with the statistics f3(present-day indigenous groups Brazil, ancient Brazilians; Mbuti). b, Same statistics as in a but using the Human Origins dataset. In both heat maps, warmer colours represent higher genetic affinities while cooler colours represent lower genetic affinities. The dot colours indicate the languages of the tested present-day populations, as shown in the bottom-left legend. |
|
|
|
Post by Admin on Dec 19, 2023 5:01:38 GMT
The analysis of the stable isotope ratio 87Sr/86Sr in the tooth enamel of JabuticabeiraII_102_1300BP (0.7111) also points at a different provenance for this female individual, possibly from another coastal location, when compared with the JabuticabeiraII_~2400BP group (0.7095 ± 0.000096; n = 7) (Supplementary Table 16). This could also indicate a dietary change, since a mixed marine and C3-resource diet has already been described for JabuticabeiraII_102_1300BP, in contrast to the high marine protein intake of older individuals74. Instead, the absence of a distinctive Jê-related signal in GalhetaIV_1200BP, considered to be the typical Jê site on the coast, points at a certain level of demic continuity with Sambaqui groups after the arrival of ceramics and the end of shellmound construction. Therefore, it suggests that cultural diffusion might have also been an important mechanism in the spread of ceramics across the Atlantic coast of Brazil, as indicated by previous studies2,21,23,75. With the Human Origins dataset, we first expanded the previous findings using f3 outgroup statistics (Fig. 3b). Moreover, f4 statistics of the form f4(Mbuti, Brazilian ancient groups; present-day indigenous groups—left, present-day indigenous groups—right) revealed that all Sambaqui individuals show a significant genetic attraction to the Xavánte (Jê-speaking) in contrast to the other available indigenous populations (Fig. 4). To investigate whether the influence of Jê-related ancestry in the Sambaqui individuals from the southern coast can be attributed specifically to either Kaingang or Xavánte, we performed the test f4(Tanzania_3000BP, Sambaqui groups; Xavánte, Kaingang_burial_100BP) (Supplementary Data 5). Here we used ancient African individuals73 to mitigate biases due to attraction between ancient DNA samples. Our results show that all Jabuticabeira II individuals are equally associated with both tested sources of Jê ancestry (Kaingang and Xavánte) (|Z| < 1.71). This suggests that the specific Jê-related ancestry contributing to southern Sambaqui groups is missing in our ancient and present-day genetic dataset. More genomic data from other Jê-speaking groups are needed to accurately assign a specific genetic contribution. Fig. 4: f4 statistics between ancient groups/individuals and present-day Brazilian groups. 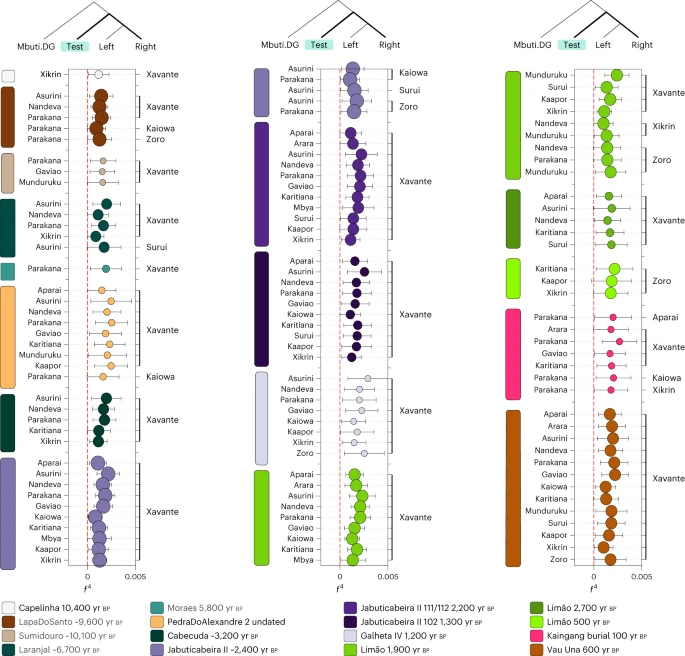 The f4 statistics polarized to positive values (Z > 3) performed on ancient Brazilian genome-wide data and present-day indigenous groups using the Human Origins dataset for f4(Mbuti, TEST; present-day indigenous groups Brazil—left, present-day indigenous group Brazil—right). The point sizes represent the number of SNPs used to compute the f4 tests (at least 20,000 SNPs). The colours correspond to the TEST ancient group/individual. The bars represent f4 statistics ± 3 standard errors. Figure related to Supplementary Data 5. |
|
|
|
Post by Admin on Dec 21, 2023 0:35:26 GMT
Links with ceramists from Amazonia and northeastern Brazil
To investigate the chronological depth of the shared ancestry between Sambaqui and Jê-, Tupi- and Karib-speaking groups, we sequenced individuals from late Holocene archaeological sites in the Cerrado of northeastern Brazil and the lower Amazon Forest. The former is associated with the Una tradition (Vau_Una_600BP), a ceramic type made by horticulturists that occupied a vast territory in central and northeastern Brazil76, and the latter is associated with the Koriabo tradition (Palmeiras Xingu_500BP), a late pre-colonial/early colonial archaeological culture (~1,200–1,600 CE) that may represent the southernmost Karib expansion in South America57,59,62,63.
The genetic patterns obtained by performing f4 tests on Vau_Una_600BP and present-day Native American populations from the Human Origins dataset show strong evidence of genetic similarities between the Una-context individual and Xavánte, when compared with Tupi (Tupi Mondé, Arikén and Tupi-Guarani) and Karib populations (Arara and Apalai) (Fig. 4). This provides direct evidence for the association of Jê-speaking populations with pottery makers of the Una tradition. The results of the f4 test performed on Brazilian indigenous populations included in the Illumina dataset show that Palmeiras Xingu_500BP shares genetic affinities with Arara, a Karib-speaking group from the lower Amazon, and with the Tupi-speaking Surui Paiter (Extended Data Fig. 5).
In comparison to all Sambaqui individuals analysed here, both Vau_Una_600BP and Palmeiras Xingu_500BP present a genetic attraction to the latest burial from the sambaqui do Limão (Limão_500BP), indicating some level of shared genetic drift in the most recent past.
The Population Y signal
We investigated the presence of the Population Y signal in the newly produced data with f4 statistics of the form f4(Mbuti, Papuan/Onge/Australian; present-day Mexicans, ancient Brazilians)65. The only ancient Brazilian group showing significant affinity to Onge, compared with present-day Mexicans, is the JabuticabeiraII_~2400BP group. The signal is mainly driven by one individual (JBT009—burial 38), but it remains for the entire group even after the exclusion of JBT009. Similarly, there is significant genetic attraction between Onge and one individual from the Cabeçuda_3200BP group (CBE004—burial 15), while all other tests do not reach values close to significance (Supplementary Data 7). However, no evidence of the Population Y signal is found in the recent Amazonian individual Palmeiras Xingu_500BP, despite the fact that this ancestry was first described in present-day Amazonian populations; or in Capelinha_10400BP, despite its association with the paleoamerican cranial morphology10,65. We further tested the presence of differential affinity of ancient Brazilian individuals to present-day Papuans, Onge and Australians, as well as the 40,000-year-old Tianyuan genome-wide data from China77 using f4 statistics of the form f4(Mbuti, Papuan/Onge/Australian/Tianyuan; Ancient Brazilian A, Ancient Brazilian B). Only the JabuticabeiraII_~2400BP group reaches significant attraction to both Onge and Papuans, and only in comparison to LapaDoSanto_9600BP (ref. 63). This suggests either that the Population Y signal is equally widespread in most tested ancient individuals from Brazil or that previously reported attractions to non-American ancestries62,77,78 are exacerbated by the use of present-day Mexican populations in comparison to ancient groups (Supplementary Data 7).
Uniparental markers, genetic diversity and runs of homozygosity
All males in our dataset belong to Y-chromosome haplogroup Q1b, which has the highest frequency in present-day South Americans. To the limit of the available SNP coverage, the male individuals from Jabuticabeira II carry either the common haplogroup Q1b1a1a-M3 or the currently rare haplogroup Q1a2a1b-CTS1780, confirming its higher frequency in ancient South Americans63 (Supplementary Data 1).
The mtDNA analysis shows that all newly studied individuals belong to American-specific mtDNA haplogroups (A2, B2, C1b, C1c, C1d1 and D1) (Supplementary Data 1). An exception is individual Loca Do Suin_9100BP, who carries the extremely rare and primarily North American mtDNA haplogroup C4c. Finding this mtDNA lineage in Brazil during the early Holocene provides additional support to the possibility that haplogroup C4c entered the Americas during early peopling events79. On the basis of the mtDNA diversity, we tested the presence of sub-structure among Sambaqui groups. Our results show a level of differentiation between Sambaqui individuals from the south coast and those from the southeast coast (Extended Data Fig. 6).
At the Jabuticabeira II site, 16 individuals share the same mtDNA haplogroup C1c with a maximum of one nucleotide distance among all mtDNA sequences. The only exception is represented by JabuticabeiraII_102_1300BP, who carries mtDNA haplogroup B2 (ref. 63). This pattern of uniparental markers, considered alongside the generally low pairwise mismatch rate, could be compatible with a scenario of consanguinity among Jabuticabeira II individuals (Extended Data Fig. 7). To test this, we calculated runs of homozygosity (ROH)80. Those results revealed a large number of short ROH (4–8 cM) in the JabuticabeiraII_~2400BP group, suggesting a smaller effective population size (2n of ~400 to ~1,600 individuals contributing to the next generation) than for younger burials from the same site (Fig. 5a). Therefore, rather than recent consanguinity, this genetic pattern is consistent with a bottlenecked population and calls into question the expectation of large demography in Sambaqui societies. Studies of pre-contact subsistence fisheries using data from the Cubatão I site have also indicated a lower-than-expected population size among southern Sambaqui groups81. Contemporaneous individuals from the sambaqui do Limão present a similar ROH profile, while the Limao_500BP individual shows a pattern consistent with first-cousin consanguinity (Fig. 5a).
|
|
|
|
Post by Admin on Dec 22, 2023 18:29:44 GMT
Fig. 5: ROH profiles and heterozygosity of the ancient coastal groups. 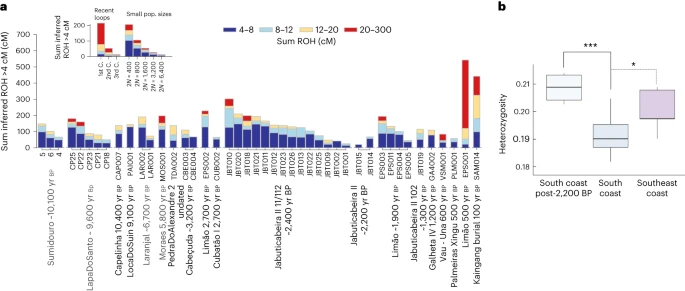 a, Sum of ROH fragments higher than 4 cM for each individual with more than 190,000 SNPs sorted by population name and in chronological order. The insert provides a legend of individual ROH profiles for recent loops (parents from 1st to 3rd cousins (C.)) and small population sizes. b, Heterozygosity distribution among the tested groups. This was calculated on the basis of the pseudo-diploid genotypes of three Sambaqui groups: south coast (n = 17), southeast coast (n = 5) and 2,200–1,200 yr BP individuals from the south coast (n = 4). In the box plots, the central line represents the median, the box edges represent the 25th and 75th percentiles, and the whiskers show the distribution of the remaining variation. The connectors mark the significant results obtained with the non-parametric Kruskal–Wallis test (P = 0.001), followed by the post hoc Conover’s test for multiple comparisons using the false discovery rate correction method (*P = 0.01914; ***P = 0.00089). Finally, the south coast Sambaqui groups (JabuticabeiraII_~2400BP, Cabeçuda_3200BP and CubatãoI_~2700BP) show lower heterozygosity levels than those at the southeast coast site (sambaqui do Limão) and even lower than late Sambaqui individuals from the south coast (JabuticabeiraII_111/112_~2200BP, JabuticabeiraII_102_1300BP and GalhetaIV_1200BP) (Fig. 5b). The increase in heterozygosity through time in southern Sambaqui groups is probably associated with gene flow of Jê-related ancestry from the inland detected here by 2,200 yr BP. |
|


 Catarina and Espírito Santo states, respectively) do not constitute a genetically homogenous population, as previously suggested by the analyses of cranial and dental morphological variation21,23.
Catarina and Espírito Santo states, respectively) do not constitute a genetically homogenous population, as previously suggested by the analyses of cranial and dental morphological variation21,23.