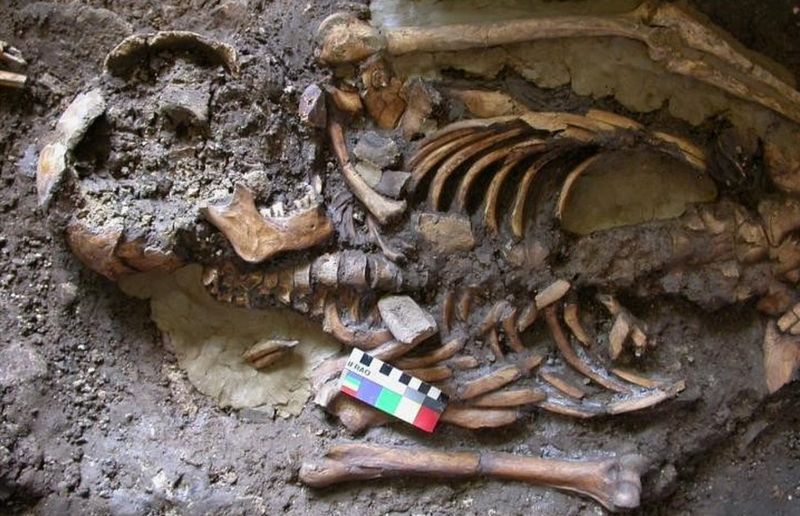
Post by Admin on May 1, 2021 23:30:27 GMT
Supplementary Note 6
Phenotypes of interest
To get a picture of the phenotypic characteristics of our samples, we examined genes which
have been associated with particular phenotypes in modern populations, including some loci
which have been subject to selection in European populations. To investigate skin tone in our
samples we began by using the 8-plex prediction model66, a tool developed for forensic
applications. We found the skin colour results for Kotias, Satsurblia and Bichon to be
inconclusive (Supplementary Table 20). To explore further we looked at genotypes in two
pigmentation genes proposed to have been strongly selected in the ancestors of modern
Europeans, namely SLC45A2 and SLC24A514,67–71. Skin colour tends to get progressively paler
with increasing distance from the equator72 and this pattern is thought be the result of natural
selection. In higher latitudes, with restricted ultraviolet radiation exposure, lighter skin colour
confers a selective advantage as it allows increased dermal vitamin D synthesis73,74. Selected
SNPs in the SLC45A2 and SLC24A5 genes (rs16891982 and rs1426654 respectively)
contribute to lightening of skin and are almost fixed in modern Europeans14,67–71. We found that
Kotias and Satsurblia have the ancestral version of the SLC45A2 (rs16891982) variant but both
CHG have the selected version of the SLC24A5 (rs1426654) gene (Supplementary Table 21)
encompassed by the most commonly associated haplotype (C11) found in modern
populations14. Bichon on the other hand has the ancestral version of both genes suggesting that
our Caucasus hunter-gatherers may have had lighter skin than our western hunter-gatherer,
Bichon (Supplementary Table 20).
We used the Hirisplex online tool to predict hair and eye colour for our samples75. We found it
most likely that Bichon, Kotias and Satsurblia had dark/black hair and brown eyes
(Supplementary Tables 22 & 23). It is unlikely that any of our samples were able to drink milk
into adulthood as all samples had the ancestral genotype at two positions (rs4988235 and
rs182549) upstream of the LCT locus where the derived genotype is associated in Europeans
with the ability to process lactose. The ability to digest milk is thought to have been driven to
high frequencies in Europe subsequent to the introduction of farming2,76
Supplementary Note 7
Runs of homozygosity
To gain an insight into past population structure we examined runs of homozygosity (ROH) in
our ancient samples. ROH occur when identical extended regions of the genome are inherited
from both parents and their distribution can be informative about past population
demography2,60–62. Long homozygous genomic stretches provide evidence for recent endogamy
because recombination has not acted to break down these long tracts which are identical by
descent. In contrast short runs can be indicative of an ancient population bottleneck. After such
a constrictive event a population will experience a period of increased inbreeding creating long
homozygous haplotypes but these segments can be broken up by recombination over time as
the population expands, creating short homozygous runs.
Examination of ROH requires dense diploid genotypes. We used imputation to maximise the
information content of our most ancient sample, Satsurblia, which was sequenced to 1.44x.
Imputation allows the inference of missing genotypes by comparing surrounding haplotypes in
the sample to those found in a phased reference panel and has been shown to be a valid
method for leveraging palaeogenomic data2. We were concerned that the haplotypes present in
Satsurblia may not be well represented by haplogroups in our modern dataset. To test if CHG
genotypes could be accurately imputed we down-sampled our high coverage genome Kotias to
~1x and compared 546,625 imputed genotypes which overlapped with our confidently called
high coverage genotypes for the filtered Human Origins dataset (Supplementary Fig. 10). When
we imposed a genotype probability of 0.99 we found that 85% of loci were retained and of those
99.41% (97.91% of heterozygotes) matched our high coverage calls (Supplementary Fig. 8).
This high concordance rate supports the use of imputed CHG data in our analyses.
ROH analysis was carried out as described in the methods section. When we plotted short ROH
(<1.6 Mb) against long ROH (≥ 1.6 Mb) (Figure 3A)2,61 we found that our three hunter-gatherer
samples, Satsurblia, Kotias and Bichon locate with other hunter-gatherer samples in a region of
the plot with a relative excess of both short and long ROH compared to Neolithic farmers. These
short runs suggest an ancestrally restricted population size for CHG as well as WHG which
could reflect the effect of a reduced population size in glacial refugia. This contrasts with the
relatively low frequency ROH found for Neolithic samples, perhaps because the ancestors of
these people resided in a location further south with a more moderate climate during the LGM,
permissive of a larger effective population size. Longer runs of ROH found in hunter-gatherer
samples are compatible with more recent consanguinity in their family lines than that
experienced by Neolithic farmers.
Kotias, Bichon and Loschbour all overlap with individuals from America with the former two
samples also overlapping Oceanic individuals. Both American and Oceanic populations have
experienced a population bottleneck during their histories60. Of all the ancient samples
Satsurblia has the most long ROH and lies closest to the Onge, indigenous people from the
Andaman islands. This island population has experienced long term isolation resulting in a small
ancestral population size63 and recent population reduction after colonisation by the British in
185864.
We also placed our ROH into size bins60 (Figure 2B) and found Neolithic farmers to have a ROH
distribution that follows a similar pattern to modern Eurasian groups. In contrast huntergatherers had a relative excess of ROH <4 Mb in size, a signature of a small ancestral population size. Compared to other hunter-gatherer samples, Satsurblia has an excess of long ROH 4-16 Mb in size suggestive of a more recent interbreeding event in the family history of this individual.
Phenotypes of interest
To get a picture of the phenotypic characteristics of our samples, we examined genes which
have been associated with particular phenotypes in modern populations, including some loci
which have been subject to selection in European populations. To investigate skin tone in our
samples we began by using the 8-plex prediction model66, a tool developed for forensic
applications. We found the skin colour results for Kotias, Satsurblia and Bichon to be
inconclusive (Supplementary Table 20). To explore further we looked at genotypes in two
pigmentation genes proposed to have been strongly selected in the ancestors of modern
Europeans, namely SLC45A2 and SLC24A514,67–71. Skin colour tends to get progressively paler
with increasing distance from the equator72 and this pattern is thought be the result of natural
selection. In higher latitudes, with restricted ultraviolet radiation exposure, lighter skin colour
confers a selective advantage as it allows increased dermal vitamin D synthesis73,74. Selected
SNPs in the SLC45A2 and SLC24A5 genes (rs16891982 and rs1426654 respectively)
contribute to lightening of skin and are almost fixed in modern Europeans14,67–71. We found that
Kotias and Satsurblia have the ancestral version of the SLC45A2 (rs16891982) variant but both
CHG have the selected version of the SLC24A5 (rs1426654) gene (Supplementary Table 21)
encompassed by the most commonly associated haplotype (C11) found in modern
populations14. Bichon on the other hand has the ancestral version of both genes suggesting that
our Caucasus hunter-gatherers may have had lighter skin than our western hunter-gatherer,
Bichon (Supplementary Table 20).
We used the Hirisplex online tool to predict hair and eye colour for our samples75. We found it
most likely that Bichon, Kotias and Satsurblia had dark/black hair and brown eyes
(Supplementary Tables 22 & 23). It is unlikely that any of our samples were able to drink milk
into adulthood as all samples had the ancestral genotype at two positions (rs4988235 and
rs182549) upstream of the LCT locus where the derived genotype is associated in Europeans
with the ability to process lactose. The ability to digest milk is thought to have been driven to
high frequencies in Europe subsequent to the introduction of farming2,76
Supplementary Note 7
Runs of homozygosity
To gain an insight into past population structure we examined runs of homozygosity (ROH) in
our ancient samples. ROH occur when identical extended regions of the genome are inherited
from both parents and their distribution can be informative about past population
demography2,60–62. Long homozygous genomic stretches provide evidence for recent endogamy
because recombination has not acted to break down these long tracts which are identical by
descent. In contrast short runs can be indicative of an ancient population bottleneck. After such
a constrictive event a population will experience a period of increased inbreeding creating long
homozygous haplotypes but these segments can be broken up by recombination over time as
the population expands, creating short homozygous runs.
Examination of ROH requires dense diploid genotypes. We used imputation to maximise the
information content of our most ancient sample, Satsurblia, which was sequenced to 1.44x.
Imputation allows the inference of missing genotypes by comparing surrounding haplotypes in
the sample to those found in a phased reference panel and has been shown to be a valid
method for leveraging palaeogenomic data2. We were concerned that the haplotypes present in
Satsurblia may not be well represented by haplogroups in our modern dataset. To test if CHG
genotypes could be accurately imputed we down-sampled our high coverage genome Kotias to
~1x and compared 546,625 imputed genotypes which overlapped with our confidently called
high coverage genotypes for the filtered Human Origins dataset (Supplementary Fig. 10). When
we imposed a genotype probability of 0.99 we found that 85% of loci were retained and of those
99.41% (97.91% of heterozygotes) matched our high coverage calls (Supplementary Fig. 8).
This high concordance rate supports the use of imputed CHG data in our analyses.
ROH analysis was carried out as described in the methods section. When we plotted short ROH
(<1.6 Mb) against long ROH (≥ 1.6 Mb) (Figure 3A)2,61 we found that our three hunter-gatherer
samples, Satsurblia, Kotias and Bichon locate with other hunter-gatherer samples in a region of
the plot with a relative excess of both short and long ROH compared to Neolithic farmers. These
short runs suggest an ancestrally restricted population size for CHG as well as WHG which
could reflect the effect of a reduced population size in glacial refugia. This contrasts with the
relatively low frequency ROH found for Neolithic samples, perhaps because the ancestors of
these people resided in a location further south with a more moderate climate during the LGM,
permissive of a larger effective population size. Longer runs of ROH found in hunter-gatherer
samples are compatible with more recent consanguinity in their family lines than that
experienced by Neolithic farmers.
Kotias, Bichon and Loschbour all overlap with individuals from America with the former two
samples also overlapping Oceanic individuals. Both American and Oceanic populations have
experienced a population bottleneck during their histories60. Of all the ancient samples
Satsurblia has the most long ROH and lies closest to the Onge, indigenous people from the
Andaman islands. This island population has experienced long term isolation resulting in a small
ancestral population size63 and recent population reduction after colonisation by the British in
185864.
We also placed our ROH into size bins60 (Figure 2B) and found Neolithic farmers to have a ROH
distribution that follows a similar pattern to modern Eurasian groups. In contrast huntergatherers had a relative excess of ROH <4 Mb in size, a signature of a small ancestral population size. Compared to other hunter-gatherer samples, Satsurblia has an excess of long ROH 4-16 Mb in size suggestive of a more recent interbreeding event in the family history of this individual.