Post by Admin on Feb 19, 2023 17:15:36 GMT
Ancestry and admixture in South Asia
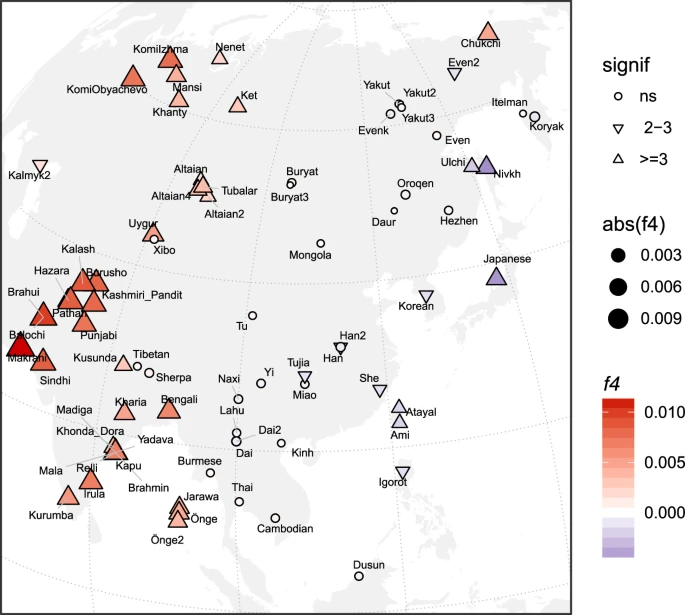
In South Asia, sampling has largely been limited to present-day populations. However, recent sampling of ancient individuals from around the Indus Valley up to 5,000 years ago [59,60] revealed two patterns. First, from PCA analyses, 5,000–4,000-year-old individuals were distributed on a cline where individuals at one end were associated with the AASI lineage and individuals at the other end were associated with basal ancestry links to ancient Iranians [59]. Narasimhan et al. [59] described the cline between individuals associated with AASI and basal Iranian ancestries the Indus Periphery cline. In a qpAdm analysis these individuals fit best as a source population for younger individuals from the Indus Periphery region and present-day South Asians [59], and thus, I denote the ancestry associated with these individuals as Indus Periphery ancestry (Box 2, ‘IP’ in Figure 2). Estimates of mixture proportions using qpAdm in 4,000–3,000-year-old individuals from northern regions of South Asia showed admixture from populations of increased Steppe ancestry associated with the Yamnaya and other Central Asian and Steppe populations [59,74]. This pattern suggests populations associated with Steppe ancestry migrated southward into the Indus Valley from 4,000 years ago, contributing to a widespread signature of Steppe ancestry across Inner Asia [59].
Reich et al. performed genome-wide sequencing of present-day Indian populations and found a north to south cline [44]. Comparison with ancient individuals from South Asia showed that all present-day Indians have a mixture of ancestry related to the AASI lineage, basal Iranian ancestry, and Steppe ancestry [59]. Northern and southern Indians are both associated with Indus Periphery ancestry observed in populations near and in the Indus Valley older than 4,000 years [59]. Southern Indian populations possess additional ancestry related to the AASI lineage beyond that found in the ancient Indus Valley individuals, which suggests that ancient individuals representing the AASI lineage, who have yet to be sampled, likely lived in southern India [59]. Northern Indians show genetic patterns similar to those found in ancient populations near the Indus Valley younger than 4,000 years; all show admixture with populations associated with Steppe ancestry [59]. These patterns illustrate that in South Asia, the formation of ancestries associated with northern and southern Indians likely post-dated 4,000 years ago, where northern Indian populations associated with the Indus Periphery cline mixed with populations of Steppe ancestry and southern Indian populations in the Indus Periphery cline primarily mixed with populations of the AASI lineage [59] (Figure 2). Just as in other regions of Asia, admixture played a key role in the formation of present-day Indian populations.
Conclusions
At the turn of the century, Cavalli-Sforza and his colleagues pushed for cataloging of genetic variation across multiple present-day human populations worldwide, including Asia, to allow deep insight into human migration patterns [2], culminating in the Human Genome Diversity Project. Prior to the availability of these datasets, Cavalli-Sforza showed how DNA sequences from humans worldwide were key to confirming modern human origins in Africa and revealing the number of dispersals into Eurasia [5]. Denser sampling in the 21st century of present-day Asians and Australasians helped to clarify that a single major dispersal was the primary contributor to all Asians and Australasians today.
While Cavalli-Sforza’s primary effort at a regional level was to characterize human genetic history in Europe [3,4], he supported similar initiatives in Asia and recognized the importance of understanding human evolutionary history in that region. Research on Asian human genetics has lagged behind that in European human genetics [96], but that has changed greatly in the last decade, with recent efforts such as the GA100K project for large-scale DNA sequencing and analysis of Asian populations [11]. In 2017, only a handful of individuals were sequenced in Asia [22], but ancient DNA studies in this region have rapidly become available since then. Sampling of ancient humans from Asia in the last five years has fundamentally changed our understanding of human movement and interaction in Asia over the last 45,000 years.
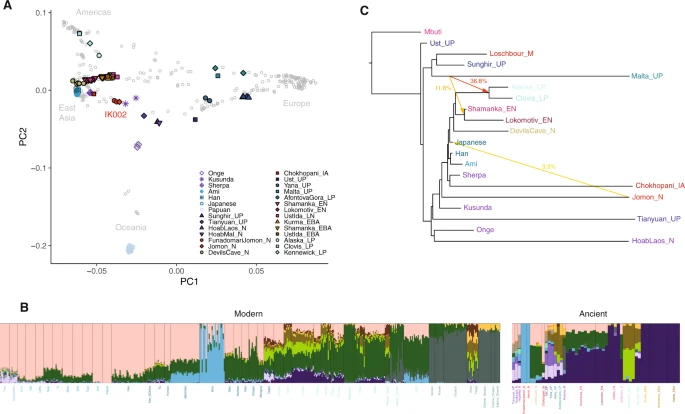
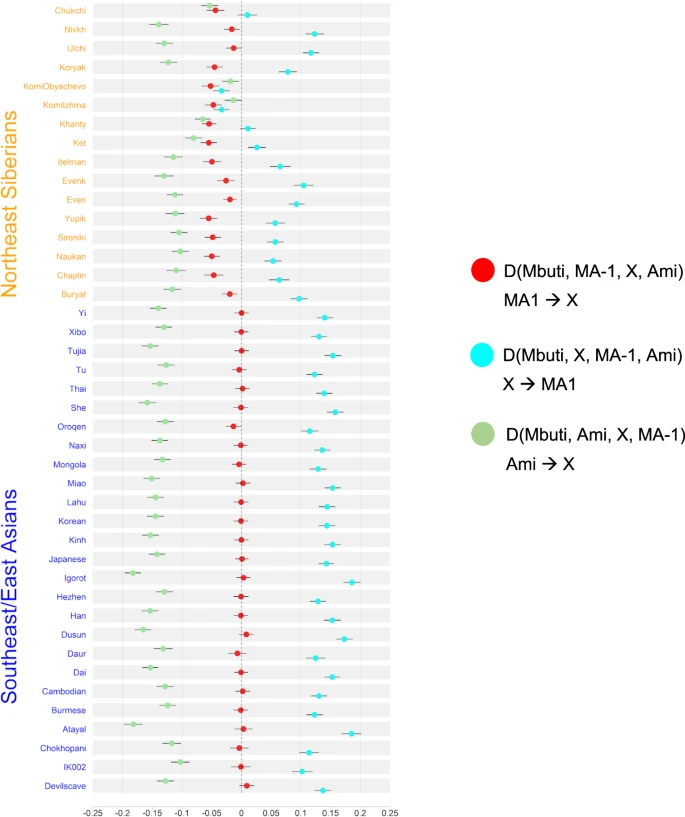
Rapid diversification of an ancestral Asian population led to at least three Asian lineages, associated with Australasians and Negritos (AA), South Asians and Andamanese Islanders (AASI), and East and Southeast Asians (ESEA). Sampling across time and space in eastern Asia indicated that the ESEA lineage was highly sub-structured in the Upper Paleolithic, and populations across Siberia, East Asia, and Southeast Asia are associated with many distinct ancestries. Today, most populations from Siberia, East Asia, and Southeast Asia are associated with a mix of ancestries sampled predominantly in ancient individuals of East Asia. Notably, deeply diverged ancestries not associated with the AA, AASI, and ESEA lineages also had a profound influence in Asia; in Upper Paleolithic Siberia, populations associated with a thriving Ancient Northern Siberian (ANS) ancestry were widespread and closely related to European hunter-gatherers (Figure 2). ANS ancestry had a lasting impact on the ancestral population contributing to the First Americans, as well as steppe populations who would later influence populations in Mongolia, Siberia, and central and southern Asia. Population movement and admixture fundamentally changed the genetic landscape for humans in the last 10,000 years (Figure 2), both because populations that once lived during the Upper Paleolithic did not continue to the present-day and because gene flow between previously isolated populations masked past genetic diversity.
With revolutionary advances in sequencing, computing, and statistics, a rich tapestry of human genetic diversity in Asia is being revealed. However, large temporal and spatial gaps still remain among the individuals sequenced. This hints at the many questions still remaining about human population history in Asia, which make it difficult to determine ancestral ranges and source populations. For instance, human movement and interaction on the Tibetan Plateau and southwest China is still unknown, and the Yangtze River region, a major contributor to rice agriculture whose populations likely influenced many regions of Asia, has yet to be sampled. Many ancestries are represented by only one or a few individuals, rendering it difficult to determine whether to attribute excess allele sharing to population structure or genetic admixture. Many demographic inference tools, like MSMC and momi2, require higher coverage and phased genomes – currently still rare for most ancient DNA studies. Greater advances in ancient DNA sequencing techniques and population genetic inference methods incorporating low coverage genomic data, along with greater systematic sampling across Asia are needed to address these limits. Despite these drawbacks, the research to date has clearly demonstrated the power of both spatial genetic sampling, as advocated by Cavalli-Sforza [1,2], and temporal genetic sampling in revealing the genetic history of humans in Asia.
www.pivotscipub.com/hpgg/2/1/0001/html
In South Asia, sampling has largely been limited to present-day populations. However, recent sampling of ancient individuals from around the Indus Valley up to 5,000 years ago [59,60] revealed two patterns. First, from PCA analyses, 5,000–4,000-year-old individuals were distributed on a cline where individuals at one end were associated with the AASI lineage and individuals at the other end were associated with basal ancestry links to ancient Iranians [59]. Narasimhan et al. [59] described the cline between individuals associated with AASI and basal Iranian ancestries the Indus Periphery cline. In a qpAdm analysis these individuals fit best as a source population for younger individuals from the Indus Periphery region and present-day South Asians [59], and thus, I denote the ancestry associated with these individuals as Indus Periphery ancestry (Box 2, ‘IP’ in Figure 2). Estimates of mixture proportions using qpAdm in 4,000–3,000-year-old individuals from northern regions of South Asia showed admixture from populations of increased Steppe ancestry associated with the Yamnaya and other Central Asian and Steppe populations [59,74]. This pattern suggests populations associated with Steppe ancestry migrated southward into the Indus Valley from 4,000 years ago, contributing to a widespread signature of Steppe ancestry across Inner Asia [59].
Reich et al. performed genome-wide sequencing of present-day Indian populations and found a north to south cline [44]. Comparison with ancient individuals from South Asia showed that all present-day Indians have a mixture of ancestry related to the AASI lineage, basal Iranian ancestry, and Steppe ancestry [59]. Northern and southern Indians are both associated with Indus Periphery ancestry observed in populations near and in the Indus Valley older than 4,000 years [59]. Southern Indian populations possess additional ancestry related to the AASI lineage beyond that found in the ancient Indus Valley individuals, which suggests that ancient individuals representing the AASI lineage, who have yet to be sampled, likely lived in southern India [59]. Northern Indians show genetic patterns similar to those found in ancient populations near the Indus Valley younger than 4,000 years; all show admixture with populations associated with Steppe ancestry [59]. These patterns illustrate that in South Asia, the formation of ancestries associated with northern and southern Indians likely post-dated 4,000 years ago, where northern Indian populations associated with the Indus Periphery cline mixed with populations of Steppe ancestry and southern Indian populations in the Indus Periphery cline primarily mixed with populations of the AASI lineage [59] (Figure 2). Just as in other regions of Asia, admixture played a key role in the formation of present-day Indian populations.
Conclusions
At the turn of the century, Cavalli-Sforza and his colleagues pushed for cataloging of genetic variation across multiple present-day human populations worldwide, including Asia, to allow deep insight into human migration patterns [2], culminating in the Human Genome Diversity Project. Prior to the availability of these datasets, Cavalli-Sforza showed how DNA sequences from humans worldwide were key to confirming modern human origins in Africa and revealing the number of dispersals into Eurasia [5]. Denser sampling in the 21st century of present-day Asians and Australasians helped to clarify that a single major dispersal was the primary contributor to all Asians and Australasians today.
While Cavalli-Sforza’s primary effort at a regional level was to characterize human genetic history in Europe [3,4], he supported similar initiatives in Asia and recognized the importance of understanding human evolutionary history in that region. Research on Asian human genetics has lagged behind that in European human genetics [96], but that has changed greatly in the last decade, with recent efforts such as the GA100K project for large-scale DNA sequencing and analysis of Asian populations [11]. In 2017, only a handful of individuals were sequenced in Asia [22], but ancient DNA studies in this region have rapidly become available since then. Sampling of ancient humans from Asia in the last five years has fundamentally changed our understanding of human movement and interaction in Asia over the last 45,000 years.
Rapid diversification of an ancestral Asian population led to at least three Asian lineages, associated with Australasians and Negritos (AA), South Asians and Andamanese Islanders (AASI), and East and Southeast Asians (ESEA). Sampling across time and space in eastern Asia indicated that the ESEA lineage was highly sub-structured in the Upper Paleolithic, and populations across Siberia, East Asia, and Southeast Asia are associated with many distinct ancestries. Today, most populations from Siberia, East Asia, and Southeast Asia are associated with a mix of ancestries sampled predominantly in ancient individuals of East Asia. Notably, deeply diverged ancestries not associated with the AA, AASI, and ESEA lineages also had a profound influence in Asia; in Upper Paleolithic Siberia, populations associated with a thriving Ancient Northern Siberian (ANS) ancestry were widespread and closely related to European hunter-gatherers (Figure 2). ANS ancestry had a lasting impact on the ancestral population contributing to the First Americans, as well as steppe populations who would later influence populations in Mongolia, Siberia, and central and southern Asia. Population movement and admixture fundamentally changed the genetic landscape for humans in the last 10,000 years (Figure 2), both because populations that once lived during the Upper Paleolithic did not continue to the present-day and because gene flow between previously isolated populations masked past genetic diversity.
With revolutionary advances in sequencing, computing, and statistics, a rich tapestry of human genetic diversity in Asia is being revealed. However, large temporal and spatial gaps still remain among the individuals sequenced. This hints at the many questions still remaining about human population history in Asia, which make it difficult to determine ancestral ranges and source populations. For instance, human movement and interaction on the Tibetan Plateau and southwest China is still unknown, and the Yangtze River region, a major contributor to rice agriculture whose populations likely influenced many regions of Asia, has yet to be sampled. Many ancestries are represented by only one or a few individuals, rendering it difficult to determine whether to attribute excess allele sharing to population structure or genetic admixture. Many demographic inference tools, like MSMC and momi2, require higher coverage and phased genomes – currently still rare for most ancient DNA studies. Greater advances in ancient DNA sequencing techniques and population genetic inference methods incorporating low coverage genomic data, along with greater systematic sampling across Asia are needed to address these limits. Despite these drawbacks, the research to date has clearly demonstrated the power of both spatial genetic sampling, as advocated by Cavalli-Sforza [1,2], and temporal genetic sampling in revealing the genetic history of humans in Asia.
www.pivotscipub.com/hpgg/2/1/0001/html