|
|
Post by Admin on Oct 26, 2023 6:37:20 GMT
Fig. 6 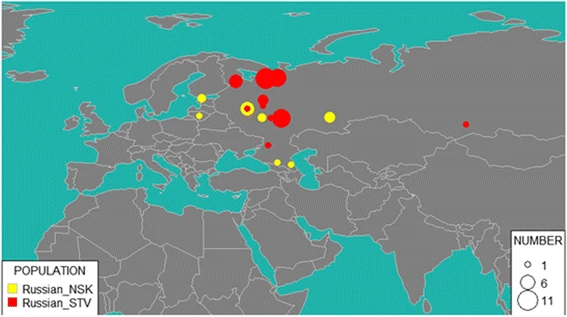 GPS results for NSK (Novosibirsk Russians) and STV (Starover Russians). Size of the bubble corresponds to the number of individuals attributed to the region Starover Russians appear to be more closely related to European Russians than Russians in Novosibirsk, with 58% identified as descendants of the migrants from various cities and villages in European part of Russia: for 45% of them, the provenance was traced to Arkhangelsk region, for 7% to Vologda region, for 2% to Yaroslavl region, and for 2% each to Tver region and Don Cossacks. Other notable ethnic component groups include 23% of Erzya and Moksha Mordva collectively, 12% of Karelian, and 5% of Veps. Altogether, GPS analysis of Starovers suggests that most of them came from northern areas of European Russia. This agrees with the slightly higher value of the Northern European component in Starovers as compared to Novosibirsk Russians. In addition to the proposed population and geographic location, the GPS algorithm also reports prediction uncertainty calculated from the distance to the nearest reference population. One of the Starover individuals was identified by GPS as a Khakas, a Turkic ethnicity living in the Republic of Khakassia located in southern Siberia, Russia. The same individual had the largest prediction uncertainty (7%) as compared to the average 3% prediction uncertainty for other Starovers samples. Typically, the prediction uncertainties which exceed 4% indicate mixed origin of an individual. For these cases, GPS algorithm should not be used. Therefore, for further analysis of Starovers and Novosibirsk individuals, we used reAdmix [39], which represents each individual as weighted sums of modern reference populations (see Fig. 7). In agreement with the GPS results, self-identified Russians from Novosibirsk appear to be more admixed than the Starovers. In Novosibirsk, 37% of genetic input came from ethnic Russians (15% from Northern Russia and 23% from Southern Russia), 25% Finno-Ugric (Veps, Karelian, Mordva), and 38% to other (Buriat, Chukchi, Chuvash, Dolgan, Evenki, Ket, Nenets, Nganasan, Selkup, Tatar, Tuvinian, Yakut, Yukaghir). Among the Starovers, 50% of ancestry was attributed to Russians (with 41% from Northern Russian and 9% of Southern Russia), 33% to Finno-Ugric (Veps, Karelian, Mordva), and 17% to other, including native Siberian populations (such as Tuva, Buryat, Yakut, Ket, Khanty). This observation supports the notion that Siberian Starovers represent relatively large heterogeneous group, which did not stay entirely isolated. |
|
|
|
Post by Admin on Oct 28, 2023 4:49:29 GMT
Fig. 7 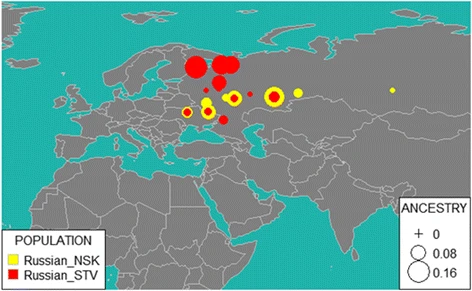 reAdmix for NSK (Novosibirsk Russians) and STV (Starover Russians). Size of the bubble corresponds to average ancestry percentage in a corresponding population Since the strict religious rules prevented Russian Starovers from marrying members of other ethnic groups, they are commonly believed to be less admixed with native Siberians than other Russian communities in the region. However, our ADMIXTURE analysis showed that the admixture profiles of Russian Starovers and Russians from Novosibirsk are similar (Fig. 2) and that both groups experienced comparable gene flow. This genetic input can be attributed to multiple known and unknown events in the history of Starovers. We can summarize our observations as follows. According to both GPS and reAdmix analyses, Starover Russians have more significant input from Northern Russians and Finno-Ugric populations than from the South of Russia. Novosibirsk Russians represent a typical mixed Russian population of the early twenty-first century; lesser degree of admixture in the genomes of Starovers point at an increase in the rates of admixture of Russian populations with neighboring ethnicities that occurred in the last 300–400 years. f3 outgroup analysis of relatedness to ancient genomes Earlier comparative studies of ancient and modern human DNA have helped to delineate human migration routes around the world [56,57,58,59,60]. We used f3 outgroup statistics [61] to test for shared genetic drift between our studied populations and selected ancient populations, namely East European hunter gatherers, Caucasus hunter gatherers, Anatolian farmers and Mal’ta (See Additional file 3: Table S1c). It was demonstrated that f3 is positive if and only if the branch supporting the population tree is longer than the two branches discordant with the population tree [62]. Therefore, large positive values of f3 show that the two tested populations had a large amount of shared population drift. All populations from the “Extended” dataset were used as test populations, with Mal’ta [9], Eastern European hunter-gatherers [56, 58], Caucasus hunter-gatherers (Jones et al. 2015) and Neolithic samples (Mathieson et al. 2015) as the reference and Yoruba as an outgroup (Additional file 3: Table S9). The summary of the findings is shown in the Fig. 8. |
|
|
|
Post by Admin on Oct 29, 2023 21:57:08 GMT
Fig. 8 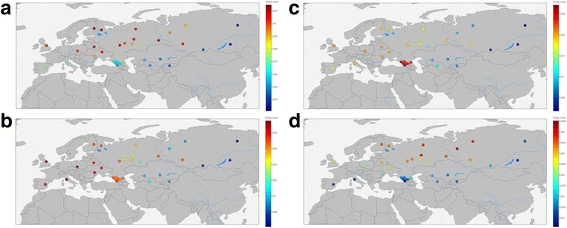 f3 values to estimate (a) Eastern European Hunter-Gatherer, b Neolithic Farmer, c Caucasus hunter-gatherer, and d) Mal’ta (Ancient North Eurasian) ancestry in modern humans This analysis confirms local inheritance of genetic structure between ancient and modern populations, as evident from consideration of aDNA samples from the Caucasus and Europe. We did not find the “source” population for our Eastern Siberian samples (Yakut and Buryat). We also confirmed that modern European population is an amalgamation of ancient European Hunter-Gatherers with Neolithic Farmers. [57, 63,64,65]. Neolithic Farmers’ genetic influence is present in a wide range of modern Eurasian populations (from the Iberian Peninsula in the West to the Altay mountains in the East). East of Altay the signal fades. Genetic signal from European Hunter Gatherers is present across several Northern Eurasian populations. The modern populations of the Caucasus show a strong signal from Caucasus hunter gatherers, that is almost absent elsewhere. Ancient North Europeans (represented by Mal’ta boy) left their genetic mark on several genomes of modern Northern Eurasians, without affecting Western or Southern Europeans or Eastern Siberians or Central Asians. |
|
|
|
Post by Admin on Nov 1, 2023 7:55:47 GMT
Discussion
Since the pioneering effort by the HapMap Consortium made in 2003 [4], multiple studies were conducted to investigate human genetic diversity, population structure, migration routes, and genotype-phenotype association [2, 8, 16, 32, 33, 55, 66,67,68,69,70,71,72,73,74,75]. These studies produced a variety of computational tools and reference datasets, leaving just a few blind spots.
One of these blind spots is in Russia, where only a handful of genome-wide human variation studies were conducted to date [10, 16, 17, 19, 23]. In this work, using the whole-genome SNP analysis, we surveyed 1019 individuals from Northern Eurasia for their genetic diversity. Newly acquired genome-wide high-density coverage for almost 30 ethnic groups in Russia enabled us to perform both inter-population and population-specific analyses. Combined with genome sequencing data available for a limited number of individuals, such as described in [23], our study provides one of the most comprehensive datasets covering genetic variation in Russia.
The relationship between genetics and geography was analysed by a combination of ADMIXTURE-based and IBD sharing approaches. We showed that Russian populations of diverse demographic histories and geographic localization share many genetic features, as reflected in their relatively tight ADMIXTURE groupings and outputs of GPS and reAdmix. The apparent positioning of some Russian samples in the genetic space of Caucasus and Siberian populations may reflect either traces of historical assimilation of these groups during the expansion of the Russians, or a recent contribution from neighbouring ethnic groups to the genomes of specific individuals. When we compared Starover and Novosibirsk Russians, representing snapshots of historical (as old as the seventeenth century) and modern (twenty-first century) Russian population, respectively, an apparent recent increase in the rates of admixture with various neighbouring population was evident. Admixture profiles of Modern Novosibirsk Russians have a lower percentage of Northern European components compared to Starovers Russians. In addition, various analyses including GPS, IBD and reAdmix suggest that Starover Russians were genetically influenced by Finno-Ugric people; this hypothesis agrees with the historical record concerning the patterns of Starover migrations within Russian Empire.
One of most curious findings involved the Bashkir, an ethnicity with an extremely complex historical background. There are three main theories describing Bashkir origins: “Turkic”, “Finno-Ugric”, and “Iranian” [76, 77]. According to the “Turkic” theory, most Bashkir genetic ancestry was formed by Turkic tribes migrating from Central Asia in the first millennium AD. The “Finno-Ugric” theory stipulates that the nucleus of Bashkir ancestry was formed by the Magyar (Hungarians), who were later assimilated by Turkic tribes and adopted a Turkic language, while the “Iranian” theory considers Bashkir to be descendants of Sarmatians from the southern Ural.
Speaking generally, our findings add weight to “Finno-Ugric” theory of the origin of Bashkir. A majority of Bashkir IBD fragments were shared with Khanty, an ethnicity related to Magyar. Interestingly, some works point out that before the thirteenth century the Hungarians were commonly called Bashkir ([78], pp. 289–294). It is surmised that the Magyar ethnicity was formed in the region between Volga and the Ural Mountains, then, at the end of the sixth century AD, moved to the Don-Kuban steppes abandoned by the Proto-Bulgarians followed by the move to their present location between Dnieper and Danube somewhat later.
Further analyses (ADMIXTURE and recent IBD) pointed to proximity of Bashkir to Turkic-speaking Tatar and Chuvash as well as to Finno-Ugric Udmurt and Khanty. In addition, results of f3 outgroup analysis indicate that Bashkir, in contrary to other Turkic speakers, were strongly influenced by Ancient Northern Eurasians, highlighting a mismatch of their cultural background and genetic ancestry and an intricacy of the historic interface between Turkic and Uralic populations. As a general pattern, the Eastern European speakers of Uralic languages share large amounts of IBD with Khanty and Ket, with Turkic speaking Bashkir being added to this rule.
It is noteworthy that the genomes of closest linguistic relatives of Bashkir, Volga Tatar, bears very little traces of East Asian or Central Siberian ancestry. Volga Tatar are a mix between Bulgar who carried a large Finno-Ugric component, Pecheneg, Kuman, Khazar, local Finno-Ugric tribes, and even Alan. Therefore, Volga Tatars are predominantly European ethnicity with a tiny contribution of East-Asian component. As most Tatar’ IBD is shared with various Turkic and Uralic populations from Volga-Ural region, an amalgamation of various cultures is evident. When the original Finno-Ugric speaking people were conquered by Turkic tribes, both Tatar and Chuvash are likely to have experience language replacement, while retaining their genetic core. Most likely, these events took place sometime around VIII century AD, after the relocation of Bulgar tribes to Volga and Kama river basins, and expansion of Turkic people.
We speculate that Bashkir, Tatar, Chuvash and Finno-Ugric speakers from Volga basin has a common Turkic component, which could have been acquired as a result of Turkic expansion to Volga-Urals region. However, the original Finno-Ugric substrate was not homogeneous: Tatar and Chuvash genomes carry mainly “Finno-Permic” component, while Bashkir carry the “Magyar” one. The fraction of the Turkic component in Bashkir is, undoubtedly, quite significant, and larger than that in Tatar and Chuvash. This component reflects the South Siberian influence on Bashkir, which makes them related to Altai, Kyrgyz, Tuvinian, and Kazakh people.
As a standalone approach, an analysis of shared IBD is not sufficient to support the Finno-Ugric hypothesis of Bashkir origin as a sole source, while pointing at temporal separation of genetic components in Bashkir. Hence, we demonstrated that Bashkir genepool is a multifaceted, multicomponent system, lacking the main “core”; it is an amalgamation of Turkic, Ugric, Finnish and Indo-European contributions. In this mosaic, it is impossible to identify the leading element. Therefore, Bashkir are the most genetically diverse ethnic group of the Volga-Urals region.
Many Siberian populations share an unusually high amount of IBD, which may be explained by a combination of the following factors: 1) shared origin, 2) relative isolation from outside world, 3) rapid recent population growth and strong founder effect in Yakut, Buryat, and Tuva, or 4) gene flow facilitated by some migrating population. The structure of these population also reflects the role of multiple South-North travel routes along the great waterways of Ob, Yenisei, and Lena, while the Siberian taiga, which is notoriously hard to traverse, to some degree prevented lateral access. On the other hand, Southern Siberia, where the steppes border the forests, is easier to travel. The same is true for the Northern Siberia, where the cold, flat tundra is suitable for travel by deer herders. These geographical limitations corralled the East-West migration to either “northern” or “southern” corridors and North-South migrations to the banks of great Siberian rivers. The footprints of these geographical restrictions could be seen in the patterns of IBD sharing between the Siberian populations studied. We christened it as the “Siberian genetic vortex”.
High IBD between West Siberian Ket and Khanty populations may reflect their relatively recent admixture with Selkup. Close genetic relationships between Ket and Tuvan can be explained by the existence of an ancient pre-Turkic and pre-Samoyedic Yenisei substrate which constitutes the main genetic component in Ket and still present in Tuva due to assimilation of extinct Yeniseian peoples (such as Kott, Arin, and Pumpokol) [79] inhabited Yenisei source area in the Southern Siberia [80].
High levels of shared IBD blocks in Altaic-speaking populations from Southern Siberia (Tuva, Buryat), North Asia (Yakut) and Central Asia (Kyrgyz) supports their recently formed common genetic core, which is geographically related to the Altay-Sayan Mountains region in Southern Siberia. Yakut and Kyrgyz populations which are now distant from this region were resettled from Southern Siberia relatively recently. It is accepted that ancestors of Yakuts (Kurykan) migrated from the Southern Yenisei to Lake Baikal area in seventh century AD, and then travelled the Lena river North in 12th -14th centuries AD [81], while Kyrgyz, who until recently were known as Yenisei Kyrgyz, migrated from Southern Siberia to Central Asia in 13th - 15th centuries AD after the collapse of the Mongol Empire [82].
The discovery of long runs of homozygosity in native Siberian populations (such as Tuva, Buryat, Yakut, Ket, Khanty) supports the earlier finding of pronounced founder effects and low genetic diversity in Siberians due to genetic drift, isolation by distance and recent population expansion events, that were made using the Y-chromosome analysis [83,84,85,86,87,88,89].
Comparative analysis of modern and ancient genomes suggests that Western Siberians have more Ancient North European ancestry (represented by Mal’ta) than other populations of the Russian Federation. Other studied populations show genetic affinity to various ancient genomes, either co-located with modern inhabitants, pointing to direct gene flow and relatively sessile population, or geographically removed, pointing to their migration to currently occupied locations.
We see that the shared genetic drift associated with hunter gatherers (Fig. 8) is correlated with Northern European ancestry of studied individuals. At the same time, the shared genetic drift of farmers has a pronounced gradient: it is large in the areas suitable for agriculture and drops to zero in Ket and Khanty-inhabited boreal forest areas of Siberia, where the climate is harsh and summers are too short for a sustainable harvest. In Siberian forests, the signal of Neolithic ancestry is no longer detected, but the ancient northern Eurasian (ANE) signal predominates instead. Possibly, the ancient Northern Eurasians met with more western groups of ancient hunters or with ancient farmers in the steppe, formed a certain population resembling the steppe samples of Yamnaya and Afanasyevo cultures, which then spread this North Eurasian component across and beyond the boreal forests of Siberia. This suggests an extensive westward migration from the steppe, discussed in detail elsewhere [56]. It is also possible that there was wave of northern or western Europeans migrating to the steppes from an opposite direction.
|
|
|
|
Post by Admin on Nov 2, 2023 18:10:34 GMT
Conclusions Our project has filled an important lacuna in the genetic map of Eurasia. We revealed the complexity of genetic structure of Northern Eurasians, the existence of East-West and North-South genetic gradients, and varying inputs of ancient populations into modern populations. In particular, we have collected evidence in support of Finno-Ugric influence on the formation of Bashkir, shed light onto the genetic make-up of Russian Starovers (Old Believers), and postulated the existence of a Great Siberian Vortex directing genetic exchanges in populations across the Siberian part of Asia. Acknowledgments We thank Prof. Roger Jelliffe for careful reading of the manuscript and Prof. Mikhail Kovalchuk for comprehensive assistance with the work. Funding TVT, PT, MT were supported by grants from National Institute of General Medical Sciences (GM068968), Eunice Kennedy Shriver National Institute of Child Health and Human Development (HD070996), National Science Foundation Division of Evolutionary Biology (1456634), and National Science Foundation Small Business Technology Transfer (1622840). TVT and OB were supported by Russian Scientific Foundation (17–14-01345). AB was supported by National Science Foundation Small Business Technology Transfer (1622840). VS was supported by Russian Scientific Foundation (16–15-00020), Russian Foundation for Basic Research (15–04-02442). VK was supported by Russian Foundation for Basic Research (16–34-60,222) and Grant of President of Russian Federation (MD-8886.2016.4). Publication costs were funded by the authors themselves. Availability of data and materials The genotype data (core dataset) are available (after the paper is published) from www.RussianGenome.ru. |
|

