Holocene transformations across Siberia and Beringia
Recent genetic studies have documented additional, Holocene-age migrations and gene flow
across the (now) Bering Strait involving both Paleoeskimos and Neoeskimos. Our genomic
data provide further insights into the peoples involved, as well as the timing of these events.
The 4 kya Saqqaq individual from Greenland12, representing Paleoeskimos, clusters with
Kolyma1, but shows greater affinity to East Asians (Figure 1; Extended Data Table 2).
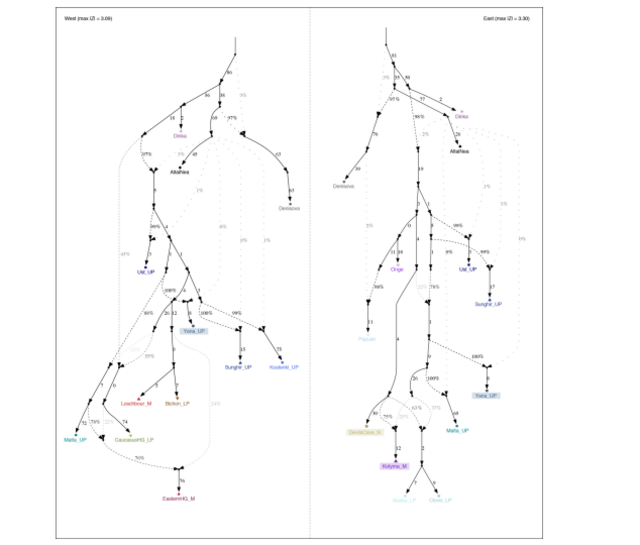
Modelling Saqqaq as a mixture of Kolyma1 and Devil’s Gate Cave (DGC), we estimate it
harbours around 20% DGC-related ancestry (Extended Data Fig. 7; Supplementary
Information 6; Supplementary Data Table 3). Individuals from the Uelen and Ekven
NeoEskimo sites (2.7 – 1.6 kya), located on the Siberian shore of the Bering Sea, closely
resemble contemporary Inuit (Figure 1, Extended Data Fig. 6). We successfully fit them as a
mixture of 69% AP (Kolyma1) and 31% Native American (Clovis) ancestry, thereby
documenting a ‘reverse’ gene flow across the Bering Sea from northwestern North America
to northeastern Siberia, in accordance with the linguistic back-migration of Eskimo-Aleut
(Extended Data Table 2; Supplementary Information 6, 9; Supplementary Data Table 1). The
source population of this gene flow post-dates the divergence of USR1 from other Native
Americans (~20.9 kya41), as the Iron Age individuals at Ekven share more alleles with
ancient Native Americans (Anzick-1, Kennewick) than with ancient Beringians (USR1),
confirming previous results from present-day Inuit49 (Extended Data Table 2). To investigate
the time of gene flow we performed linkage-disequilibrium (LD) based admixture dating50,
using Saqqaq and Anzick-1 as source populations. We find a significant weighted LD curve
with an estimated admixture date between 100 – 200 generations ago, depending on the data
set (Supplementary Information 6). While these estimates show a considerable margin of
error due to small sample size and limited amount of data for the Ekven population, they
nevertheless suggest that gene flow related to Native Americans occurred back from the
Americas into Siberia well after the disappearance of Beringia, but possibly as early as ~5
kya (~ 100 generations prior to the age of the earliest individual from Uelen and Ekven). That
process contributed to a major ancestry component of contemporary Inuit populations. A
genetic link has also been observed between North American populations speaking Na-Dene
languages (Athabascans) and Siberian populations51. It has been suggested that this link
reflects gene flow from a Paleoeskimo source represented by Saqqaq52, but a more recent
study found evidence for a ghost source population more closely related to Koryaks41. Both
admixture graph modelling (Supplementary Information 6) and chromosome-painting
symmetry tests (Extended Data Fig. 5) show that Kolyma1 is a better proxy for this ghost
ancestry than Saqqaq, therefore providing additional evidence against a contribution via a
more recent migration of Paleoeskimos.

Figure 4. Genetic legacy of ancient Eurasians. a, World-wide map of top haplotype
donations inferred by chromopainter. Each coloured symbol represents a modern recipient
population, with the colour and shape indicating the donor population contributing the
highest fraction of haplotypes to that recipient population. Geographic locations of donor
populations used in this analysis (modern Africans and ancient Eurasians) are indicated by
the corresponding larger symbols with black outline added. Extended regions of shared top
donors are visualized by spatial interpolation of the respective donor population color. b,
Major hypothesized migrations into northeast Siberia. Arrows indicate putative migrations
giving rise to Ancient North Siberians (left), Ancient Paleosiberians (middle; possible ANS
admixture scenarios indicated by small blue arrows) and Neosiberians (right). Key sample
locations for the respective time slice are indicated with symbols.
The Holocene archaeological record of northeast Siberia is marked by further changes in
material culture. We used a temporal transect of ancient Siberians from ~6 kya to 500 years
ago to investigate whether these cultural transitions were associated with genetic changes.
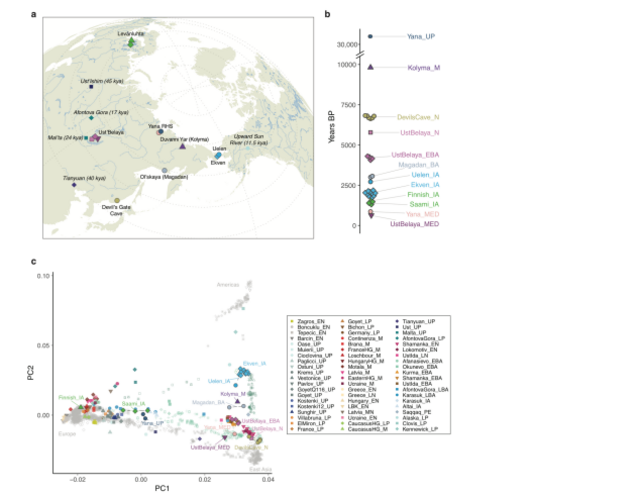
We find that in a PCA of modern non-African populations, most contemporary Siberian
populations are arranged along two separate genetic clines. The majority of individuals (here
referred to as “Neosiberians”) fall on an East-West cline stretched out along PC1 between
European populations at one end, and East Asian individuals including the ancient
individuals from Devil’s Gate Cave at the other (Figure 1). A secondary cline between East
Asians and Native Americans along PC2 includes Paleosiberian speakers and Inuit
populations. Although AP ancestry (Kolyma1) was still common in other Siberian regions
during the early Bronze Age (Extended Data Fig. 7), by the late Bronze Age we find it was
largely restricted to the northeast, exemplified by a 3 kya individual from Ol’skaya
(Magadan) that closely resembles present-day Koryaks and Itelmens. Using modern Even
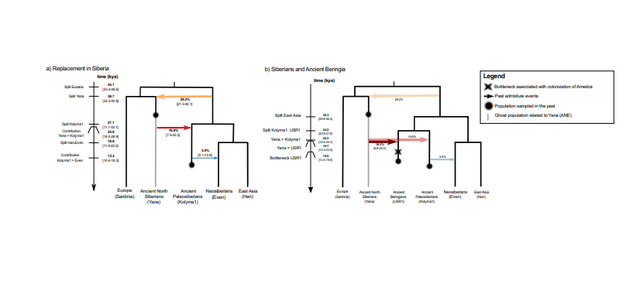
individuals to represent Neosiberians in our demographic model, we find evidence for a
recent divergence from East Asians ~13 kya, with only low levels (~6%) of AP gene flow at
~11 kya (Figure 3; Supplementary Information 7). Thus, our data provides evidence for a
second major population turnover in northeastern Siberia, with Neosiberians arriving from
the south largely replacing AP, a pattern that is also evident in chromosome painting analyses
of modern populations (Figure 4). Notable exceptions are populations such as the Ket, an
isolated population that speaks a Yeniseian language and which has previously been
described as rich in ANE-ancestry and with genetic links to Paleoeskimos51. The Ket fall on a
secondary cline parallel to Neosiberians in the chromosome painting analysis and carry ~40%
of AP ancestry (Figure 4; Extended Data Fig. 6). Our findings therefore reconcile the
proposed linguistic link between the Yeniseian speaking Ket and Na-Dene speaking
Athabascan populations (Supplementary Information 9), through shared ancestry with an AP
metapopulation that at one time was more widespread across Northern Eurasia.
Our Holocene transect reveals additional complexity in recent times, with evidence for
further episodes of gene flow and localized population replacements. A striking example is
found in the Lake Baikal region in southern Siberia, where the newly reported genomes from
Ust’Belaya and recently published neighbouring Neolithic and Bronze Age sites show a
succession of three distinct genetic ancestries over a ~6 ky time span. The earliest individuals
show predominantly East Asian ancestry, closely related to the ancient individuals from DGC
(Figure 1; Extended Data Fig. 6, 7). In the early Bronze Age (BA), we observe a resurgence
of AP ancestry (up to ~50% ancestry fraction), as well as influence of West Eurasian Steppe
ANE ancestry represented by the early BA individuals from Afanasievo in the Altai region
(~10%) (Extended Data Fig. 7; Supplementary Data Table 3). This is consistent with
previous reports of gene flow from an unknown ANE-related source into Lake Baikal huntergatherers42.
Our results suggest a southward expansion of AP as a possible source, which is
also consistent with the replacement of Y chromosome lineages observed at Lake Baikal,
from predominantly haplogroup N in the Neolithic to haplogroup Q in the BA42. Finally, the
most recent individual from Ust’Belaya, dated to ~600 years ago, falls along the Neosiberian
cline, similar to the ~760 year-old ‘Young Yana’ individual from northeastern Siberia,
demonstrating the widespread distribution of Neosiberian ancestry in the most recent epoch.
We show that most populations on this cline can be modelled as predominantly East Asian,
with varying proportions of West Eurasian ANE ancestry, with the largest proportions
observed in later ancient and modern Altaian populations (Extended Data Fig. 7;
Supplementary Data Table 3). Together, these findings show that by Holocene times there
was considerable population movement and admixture throughout southern and eastern
Siberia, with groups dispersing in multiple directions yet without clear evidence of the
wholesale population replacement seen in earlier Pleistocene times.
We further investigated whether these processes of population flux were a more widespread
phenomenon across all of Northern Eurasia. The striking spatial pattern of AP and East Asian
ancestry in modern populations observed using chromosome painting (Fig. 4) strongly
suggests that AP ancestry was once widespread, likely as far west as the Urals. At the western
edge of northern Eurasia, genetic and strontium isotope data from ancient individuals at the
Levänluhta site (Supplementary Information 1) documents the presence of Saami ancestry in
Southern Finland in the Late Holocene 1.5 kya. This ancestry component is currently limited
to the northern fringes of the region, mirroring the pattern observed for AP ancestry in
northeastern Siberia. However, while the ancient Saami individuals harbour East Asian
ancestry, we find that this is better modelled by DGC rather than AP, suggesting that AP
influence was likely restricted to the eastern side of the Urals (Extended Data Fig. 7;
Supplementary Data Table 3). Comparison of ancient Finns and Saami with their present-day
counterparts reveals additional gene flow over the past 1.6 kya, with evidence for West
Eurasian admixture into modern Saami. The ancient Finn from Levänluhta shows lower
Siberian ancestry than modern Finns (Extended Data Table 2), therefore likely representing
the dual-origin (Uralic/Scandinavian) gene pool of Finns.