Post by Admin on May 8, 2017 19:24:04 GMT
Geographic distributions of haplotypes
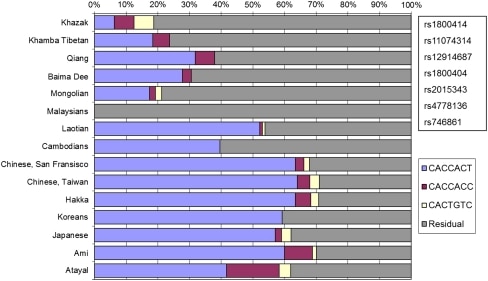
The distributions of the blue-eye associated alleles at the three haplotyped systems are presented in Fig. 2, each haplotype in contour plots, and all three grouped by population in a histogram. The actual frequencies are presented in supplemental material and in ALFRED. The alleles associated with blue eyes at all three BEH blue-eye associated haplotypes have their highest frequencies in Northwestern Europe, and the TG allele at BEH2 is essentially observed only in Europe; the ACA allele of BEH1 and the CA allele at BEH3 are at their highest frequencies in Europe, particularly in Northern and Western Europe, and have much lower frequencies elsewhere. In most of Central and East Asia, these alleles have frequencies of <20% but reach frequencies of 40% and higher in the Americas.

Fig. 3
Global distribution of the derived allele (T) of rs1800407. This figure shows the derived-allele frequencies of rs1800407.
Geographic distribution of the derived allele of rs1800407
The derived allele of rs1800407 is relatively rare compared to the blue-eye associated alleles of the three BEHs. The derived allele frequencies of rs1800407 are presented in Fig. 3. The derived allele is mostly restricted to Europe (0–11%), Southwest Asia (0–9.4%), and Central Asia (0–9.3%). Outside of this region, the derived allele is found in African Americans (1.7%), San Francisco Chinese (0.9%), the Arizona Pima (1.0%), and the Maya (3.9%).
The T allele of rs1800407 has also been associated with blue-eye penetrance (Sturm et al. 2008). We estimated haplotype frequencies for haplotypes containing rs1800407 and the three BEHs (supplemental Fig. 1). The first observation is that the blue-eye associated alleles of the three BEHs are much more common than the derived allele of rs1800407. At BEH1, the T allele of rs1800407 most commonly occurs with the AAA allele and not the ACA allele that has been associated with blue eyes. The T allele with the ACA blue-eye associated allele is the second most common combination. Other combinations occur but they are rare. The T allele of rs1800407, when seen, is commonly paired with the blue-eye associated TG allele at BEH2 only in Northern and Eastern Europeans. This association may explain the increased blue-eye penetrance seen by Sturm et al. (2008) as a type of ascertainment effect. Elsewhere the T allele is more likely to be found paired with the CA allele. We see a similar pattern at BEH3 as we see at BEH2. The blue-eye associated CA allele of BEH3 commonly pairs with the T allele only in Northwestern and Eastern Europe and the TG allele is its most common partner elsewhere.
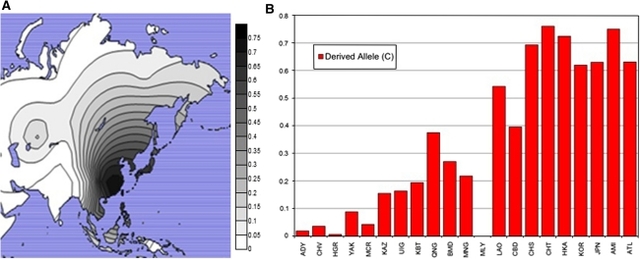
Fig. 4
Global rs1800414 derived-allele distribution and frequencies. This figure shows the distribution of the derived allele of rs1800414 interpolated on a world map (a) and as a bar graph (b).
Geographic distribution of the derived allele of rs1800414
Our data confirm that the putative light skin allele of rs1800414 (C) is found almost exclusively in East and Southeast Asia, at frequencies ranging from 0 to 76% (Fig. 4) at higher levels in eastern East Asia (62–76.1%) compared with Southeast Asia (0–54.3%) and Western China (15.5–37.5%). Outside of East and Southeast Asia, the C allele is only found in low frequencies in the Adygei, Chuvash, and Hungarians in Europe (>1–3.6%), the Yakut in Siberia (8.8%), and the Micronesians in the Pacific Islands (4.2%).
The distributions of the blue-eye associated alleles at the three haplotyped systems are presented in Fig. 2, each haplotype in contour plots, and all three grouped by population in a histogram. The actual frequencies are presented in supplemental material and in ALFRED. The alleles associated with blue eyes at all three BEH blue-eye associated haplotypes have their highest frequencies in Northwestern Europe, and the TG allele at BEH2 is essentially observed only in Europe; the ACA allele of BEH1 and the CA allele at BEH3 are at their highest frequencies in Europe, particularly in Northern and Western Europe, and have much lower frequencies elsewhere. In most of Central and East Asia, these alleles have frequencies of <20% but reach frequencies of 40% and higher in the Americas.
Fig. 3
Global distribution of the derived allele (T) of rs1800407. This figure shows the derived-allele frequencies of rs1800407.
Geographic distribution of the derived allele of rs1800407
The derived allele of rs1800407 is relatively rare compared to the blue-eye associated alleles of the three BEHs. The derived allele frequencies of rs1800407 are presented in Fig. 3. The derived allele is mostly restricted to Europe (0–11%), Southwest Asia (0–9.4%), and Central Asia (0–9.3%). Outside of this region, the derived allele is found in African Americans (1.7%), San Francisco Chinese (0.9%), the Arizona Pima (1.0%), and the Maya (3.9%).
The T allele of rs1800407 has also been associated with blue-eye penetrance (Sturm et al. 2008). We estimated haplotype frequencies for haplotypes containing rs1800407 and the three BEHs (supplemental Fig. 1). The first observation is that the blue-eye associated alleles of the three BEHs are much more common than the derived allele of rs1800407. At BEH1, the T allele of rs1800407 most commonly occurs with the AAA allele and not the ACA allele that has been associated with blue eyes. The T allele with the ACA blue-eye associated allele is the second most common combination. Other combinations occur but they are rare. The T allele of rs1800407, when seen, is commonly paired with the blue-eye associated TG allele at BEH2 only in Northern and Eastern Europeans. This association may explain the increased blue-eye penetrance seen by Sturm et al. (2008) as a type of ascertainment effect. Elsewhere the T allele is more likely to be found paired with the CA allele. We see a similar pattern at BEH3 as we see at BEH2. The blue-eye associated CA allele of BEH3 commonly pairs with the T allele only in Northwestern and Eastern Europe and the TG allele is its most common partner elsewhere.
Fig. 4
Global rs1800414 derived-allele distribution and frequencies. This figure shows the distribution of the derived allele of rs1800414 interpolated on a world map (a) and as a bar graph (b).
Geographic distribution of the derived allele of rs1800414
Our data confirm that the putative light skin allele of rs1800414 (C) is found almost exclusively in East and Southeast Asia, at frequencies ranging from 0 to 76% (Fig. 4) at higher levels in eastern East Asia (62–76.1%) compared with Southeast Asia (0–54.3%) and Western China (15.5–37.5%). Outside of East and Southeast Asia, the C allele is only found in low frequencies in the Adygei, Chuvash, and Hungarians in Europe (>1–3.6%), the Yakut in Siberia (8.8%), and the Micronesians in the Pacific Islands (4.2%).