Post by Admin on Apr 19, 2019 17:58:26 GMT
Eurasian Gene Flow Shaped the Genomes of Admixed Africans
Our results from the PCA and MSMC analysis showed a deviation of the admixed populations from the patterns observed in unadmixed (or less admixed) populations in the same geographical region. The MSMC analysis, in particular, showed that admixed Africans had patterns indicative of a decline in heterozygosity (increased bottleneck ∼60,000 ya), somewhat similarly to Eurasians. We tested whole-genome heterozygosity in these populations and found that it decreased in admixed Africans according to their Eurasian ancestry (Figure S4A). This decrease was not related to recent inbreeding, given that removing segments with long runs of homozygosity did not change the overall pattern. Our simulations suggest that decay in heterozygosity is expected after gene flow from a population with diversity comparable to that of Eurasians (Figures S4B and S4C). We further investigated heterozygosity in admixed Africans by assessing heterozygosity of the different ancestral segments in the Toubou genome. We found that admixed African-Eurasian segments had more heterozygosity (1.23 hets/kb) than segments of the genome where African-African haplotypes were present (1.19 hets/kb) (Figure S5). However, the Toubou genome segments with complete Eurasian ancestry (Eurasian-Eurasian) had considerably lower heterozygosity (∼0.96 hets/kb; Figure S5), leading to the genome-wide pattern of decay in heterozygosity observed in Africans with Eurasian ancestry (Figure S4).
We wanted to understand the consequence of admixture on the models that use the density of heterozygous sites to infer the demographic history of populations. We first tested whether the coalescent history estimated by MSMC was affected by a small proportion of mixture, such as the African mixture found in Greeks and Lebanese (ranging from 0% to 5%). We tested the Greek, Lebanese, CEU, and CHB (Han Chinese in Beijing, China) split times from the Yoruba and found that all populations split from the Yoruba ∼70,000–80,000 ya, implying that the low proportions of African admixture in the Greeks and Lebanese did not detectably affect the estimates of relative cross-coalescence rate (Figure S6A). We next tested the Toubou, who have ∼30% Eurasian ancestry. The Toubou appeared to split from Eurasians ∼30,000–40,000 ya, a time more recent than expected considering the African-Eurasian split 60,000–80,000 ya20 (Figure S6B). We tested other Africans in our dataset and found that the Sara, Laal speakers, and Yoruba split from Eurasians, as expected, ∼70,000–80,000 ya (Figure S6B). We then tested directly whether the Eurasian ancestry affected the relative cross-coalescence rate between the Toubou and Eurasians by masking some of the Eurasian ancestry in the Toubou. We used PCAdmix21 to estimate the ancestry along each chromosome and then used the identified Eurasian segments as a negative mask in our analysis. The split times between the Toubou and Lebanese, for example, increased by ∼15,000 years (Figure S6B), shifting the split date toward the expected African-Eurasian split time.
We found that, in addition to influencing the relative cross-coalescence rate, admixture can also inflate putative signals of positive selection. For example, using the PBS31 to detect recent positive selection that occurred in the Toubou after their divergence from the Yoruba, we found signals of selection on MCM6 (MIM: 601806) rs4988235, a variant associated with the lactase-persistence phenotype. This SNP was previously found to be under strong positive selection in Europeans, where it was probably advantageous to individuals living in pastoralist societies.34 The frequency of this variant in the Toubou is 2%, and it is absent from the sub-Saharan African and other Chadic samples (the Sara and Laal speakers) examined here. Although this SNP appears to be a candidate for selection, we suggest that it has probably drifted neutrally in the Toubou after the Eurasian gene flow: the Toubou have ∼30% Eurasian ancestry from a population similar to the Greeks, who have 13% derived alleles at rs4988235, suggesting an expectation of ∼3.9% of the derived allele simply from admixture. We similarly found in the Toubou signals at HERC2 (MIM: 605837) rs1129038 a major contributor to blue eye color in Europeans35 (Toubou derived allele frequency [DAF] = 0.014; Greek DAF = 0.33; Yoruba, Sara, and Laal DAF = 0), as well as a signal at SLC24A5 (MIM: 609802) rs1834640, a major contributor to pigmentation36 (Toubou DAF = 0.19; Greek DAF = 0.99; Yoruba, Sara, and Laal DAF = 0–0.04).

Figure 5
Neanderthal Ancestry Correlation with the African-Eurasian Admixture
Neanderthal ancestry is not expected in Africa, yet today many Africans carry Neanderthal-derived alleles. The plot shows that the Neanderthal ancestry proportion in Africans is correlated with gene flow from Eurasians. For example, knowing that today Eurasians carry ∼2% of Neanderthal ancestry, we observed that East Africans (Ethiopians) had ∼1% Neanderthal ancestry and ∼50% Eurasian ancestry. Correspondingly, Near Easterners showed a decline in Neanderthal ancestry proportional to their levels of African ancestry.
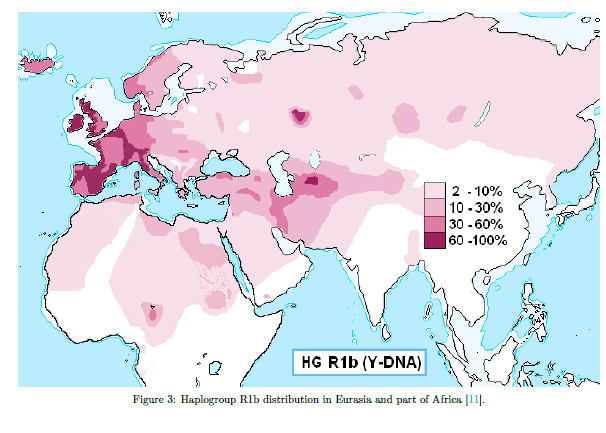
We detected the earliest Eurasian migrations to Africa in the Laal-speaking people, an isolated language group of fewer than 800 speakers who inhabit southern Chad. We estimate that mixture occurred 4,750–7,200 ya, thus after the Neolithic transition in the Near East, a period characterized by exponential growth in human population size. Environmental changes during this period (which possibly triggered the Neolithic transition) also facilitated human migrations. The African Humid Period, for example, was a humid phase across North Africa that peaked 6,000–9,000 ya37 and biogeographically connected Africa to Eurasia, facilitating human movement across these regions.38 In Chad, we found a Y chromosome lineage (R1b-V88) that we estimate emerged during the same period 5,700–7,300 ya (Figure 3B). The closest related Y chromosome groups today are widespread in Eurasia and have been previously associated with human expansions to Europe.39, 40 We estimate that the Eurasian R1b lineages initially diverged 7,300–9,400 ya, at the time of the Neolithic expansions. However, we found that the African and Eurasian R1b lineages diverged 17,900–23,000 ya, suggesting that genetic structure was already established between the groups who expanded to Europe and Africa. R1b-V88 was previously found in Central and West Africa and was associated with a mid-Holocene migration of Afro-asiatic speakers through the central Sahara into the Lake Chad Basin.8 In the populations we examined, we found R1b in the Toubou and Sara, who speak Nilo-Saharan languages, and also in the Laal people, who speak an unclassified language. This suggests that R1b penetrated Africa independently of the Afro-asiatic language spread or passed to other groups through admixture.
In addition to the early Eurasian migration to Africa ∼6,000 ya, a second migration ∼3,000 ya affected the Toubou population in northern Chad but had no detectable genetic impact on other Chadian populations. This migration appears to be associated with the previously reported Eurasian backflow into East Africa, given that the source populations and dates of mixture are similar. Occurring at the start of the Iron Age, these migrations could have been facilitated by advances in warfare and transportation technology in the Near East. It is uncertain why the impact of this migration in Chad affected only the Toubou. The African ancestral component in the Toubou is best represented by the Laal-speaking population, suggesting that the African-Eurasian mixture probably occurred in Chad. However, ethnolinguistic barriers could have already been established at this time between the Chad groups, preventing a widespread dissemination of the Eurasian ancestry. The Toubou, despite their Islamic faith, do not show the genetic admixture detected in many Near Eastern and North African populations around 1,100 ya,41 suggesting conversion without population mixing at this time. They did, however, receive additional Eurasian ancestry in the past 200 years from a source represented by North African populations such as Tunisians, Mozabite, Algerians, and Sahrawi (Figure 3C). This recent interaction could have been promoted by the nomadic lifestyle of the present-day Toubou and a shared Muslim religion with North Africans. Unsurprisingly, we also detected a likely mixing of Chad populations in the sample from the capital, which could be even more recent.
Eurasian backflow into Africa thus appears to have been a recurrent event in the history of many Africans, given its considerable impact on their genomes. Although population mixture in general is a process that increases genetic diversity, we observed a decrease in heterozygosity in the admixed Africans. Our simulations showed that these results are expected after mixture at these proportions with the Eurasians who suffered a significant bottleneck at the time of their exodus from Africa ∼60,000 ya. Consequently, we found that mixture can complicate interpretation of the coalescent history inferred from models that use the density of heterozygous sites in their implementations. In addition, we detected in admixed Africans an inflation of positive-selection signals on alleles associated with adult lactose tolerance and pigmentation in Europeans, but we suggest that these alleles have drifted neutrally in Africans after admixture. Furthermore, we detected Neanderthal ancestry in admixed Africans and found it to be proportional to their Eurasian ancestry. Similarly, in admixed Near Easterners, we found a decrease in Neanderthal ancestry proportional to the gene flow they have received from Africans. Although a higher genetic affinity of Neanderthals to Europeans than to Near Easterners was previously interpreted as additional Neanderthal admixture in the history of Europeans,42 we propose that a more parsimonious explanation for these observations is that African-Eurasian mixtures both introduced Neanderthal ancestry to Africa and “diluted” the Neanderthal ancestry in the Near East.
Am J Hum Genet. 2016 Dec 1; 99(6): 1316–1324.
Our results from the PCA and MSMC analysis showed a deviation of the admixed populations from the patterns observed in unadmixed (or less admixed) populations in the same geographical region. The MSMC analysis, in particular, showed that admixed Africans had patterns indicative of a decline in heterozygosity (increased bottleneck ∼60,000 ya), somewhat similarly to Eurasians. We tested whole-genome heterozygosity in these populations and found that it decreased in admixed Africans according to their Eurasian ancestry (Figure S4A). This decrease was not related to recent inbreeding, given that removing segments with long runs of homozygosity did not change the overall pattern. Our simulations suggest that decay in heterozygosity is expected after gene flow from a population with diversity comparable to that of Eurasians (Figures S4B and S4C). We further investigated heterozygosity in admixed Africans by assessing heterozygosity of the different ancestral segments in the Toubou genome. We found that admixed African-Eurasian segments had more heterozygosity (1.23 hets/kb) than segments of the genome where African-African haplotypes were present (1.19 hets/kb) (Figure S5). However, the Toubou genome segments with complete Eurasian ancestry (Eurasian-Eurasian) had considerably lower heterozygosity (∼0.96 hets/kb; Figure S5), leading to the genome-wide pattern of decay in heterozygosity observed in Africans with Eurasian ancestry (Figure S4).
We wanted to understand the consequence of admixture on the models that use the density of heterozygous sites to infer the demographic history of populations. We first tested whether the coalescent history estimated by MSMC was affected by a small proportion of mixture, such as the African mixture found in Greeks and Lebanese (ranging from 0% to 5%). We tested the Greek, Lebanese, CEU, and CHB (Han Chinese in Beijing, China) split times from the Yoruba and found that all populations split from the Yoruba ∼70,000–80,000 ya, implying that the low proportions of African admixture in the Greeks and Lebanese did not detectably affect the estimates of relative cross-coalescence rate (Figure S6A). We next tested the Toubou, who have ∼30% Eurasian ancestry. The Toubou appeared to split from Eurasians ∼30,000–40,000 ya, a time more recent than expected considering the African-Eurasian split 60,000–80,000 ya20 (Figure S6B). We tested other Africans in our dataset and found that the Sara, Laal speakers, and Yoruba split from Eurasians, as expected, ∼70,000–80,000 ya (Figure S6B). We then tested directly whether the Eurasian ancestry affected the relative cross-coalescence rate between the Toubou and Eurasians by masking some of the Eurasian ancestry in the Toubou. We used PCAdmix21 to estimate the ancestry along each chromosome and then used the identified Eurasian segments as a negative mask in our analysis. The split times between the Toubou and Lebanese, for example, increased by ∼15,000 years (Figure S6B), shifting the split date toward the expected African-Eurasian split time.
We found that, in addition to influencing the relative cross-coalescence rate, admixture can also inflate putative signals of positive selection. For example, using the PBS31 to detect recent positive selection that occurred in the Toubou after their divergence from the Yoruba, we found signals of selection on MCM6 (MIM: 601806) rs4988235, a variant associated with the lactase-persistence phenotype. This SNP was previously found to be under strong positive selection in Europeans, where it was probably advantageous to individuals living in pastoralist societies.34 The frequency of this variant in the Toubou is 2%, and it is absent from the sub-Saharan African and other Chadic samples (the Sara and Laal speakers) examined here. Although this SNP appears to be a candidate for selection, we suggest that it has probably drifted neutrally in the Toubou after the Eurasian gene flow: the Toubou have ∼30% Eurasian ancestry from a population similar to the Greeks, who have 13% derived alleles at rs4988235, suggesting an expectation of ∼3.9% of the derived allele simply from admixture. We similarly found in the Toubou signals at HERC2 (MIM: 605837) rs1129038 a major contributor to blue eye color in Europeans35 (Toubou derived allele frequency [DAF] = 0.014; Greek DAF = 0.33; Yoruba, Sara, and Laal DAF = 0), as well as a signal at SLC24A5 (MIM: 609802) rs1834640, a major contributor to pigmentation36 (Toubou DAF = 0.19; Greek DAF = 0.99; Yoruba, Sara, and Laal DAF = 0–0.04).
Figure 5
Neanderthal Ancestry Correlation with the African-Eurasian Admixture
Neanderthal ancestry is not expected in Africa, yet today many Africans carry Neanderthal-derived alleles. The plot shows that the Neanderthal ancestry proportion in Africans is correlated with gene flow from Eurasians. For example, knowing that today Eurasians carry ∼2% of Neanderthal ancestry, we observed that East Africans (Ethiopians) had ∼1% Neanderthal ancestry and ∼50% Eurasian ancestry. Correspondingly, Near Easterners showed a decline in Neanderthal ancestry proportional to their levels of African ancestry.
We detected the earliest Eurasian migrations to Africa in the Laal-speaking people, an isolated language group of fewer than 800 speakers who inhabit southern Chad. We estimate that mixture occurred 4,750–7,200 ya, thus after the Neolithic transition in the Near East, a period characterized by exponential growth in human population size. Environmental changes during this period (which possibly triggered the Neolithic transition) also facilitated human migrations. The African Humid Period, for example, was a humid phase across North Africa that peaked 6,000–9,000 ya37 and biogeographically connected Africa to Eurasia, facilitating human movement across these regions.38 In Chad, we found a Y chromosome lineage (R1b-V88) that we estimate emerged during the same period 5,700–7,300 ya (Figure 3B). The closest related Y chromosome groups today are widespread in Eurasia and have been previously associated with human expansions to Europe.39, 40 We estimate that the Eurasian R1b lineages initially diverged 7,300–9,400 ya, at the time of the Neolithic expansions. However, we found that the African and Eurasian R1b lineages diverged 17,900–23,000 ya, suggesting that genetic structure was already established between the groups who expanded to Europe and Africa. R1b-V88 was previously found in Central and West Africa and was associated with a mid-Holocene migration of Afro-asiatic speakers through the central Sahara into the Lake Chad Basin.8 In the populations we examined, we found R1b in the Toubou and Sara, who speak Nilo-Saharan languages, and also in the Laal people, who speak an unclassified language. This suggests that R1b penetrated Africa independently of the Afro-asiatic language spread or passed to other groups through admixture.
In addition to the early Eurasian migration to Africa ∼6,000 ya, a second migration ∼3,000 ya affected the Toubou population in northern Chad but had no detectable genetic impact on other Chadian populations. This migration appears to be associated with the previously reported Eurasian backflow into East Africa, given that the source populations and dates of mixture are similar. Occurring at the start of the Iron Age, these migrations could have been facilitated by advances in warfare and transportation technology in the Near East. It is uncertain why the impact of this migration in Chad affected only the Toubou. The African ancestral component in the Toubou is best represented by the Laal-speaking population, suggesting that the African-Eurasian mixture probably occurred in Chad. However, ethnolinguistic barriers could have already been established at this time between the Chad groups, preventing a widespread dissemination of the Eurasian ancestry. The Toubou, despite their Islamic faith, do not show the genetic admixture detected in many Near Eastern and North African populations around 1,100 ya,41 suggesting conversion without population mixing at this time. They did, however, receive additional Eurasian ancestry in the past 200 years from a source represented by North African populations such as Tunisians, Mozabite, Algerians, and Sahrawi (Figure 3C). This recent interaction could have been promoted by the nomadic lifestyle of the present-day Toubou and a shared Muslim religion with North Africans. Unsurprisingly, we also detected a likely mixing of Chad populations in the sample from the capital, which could be even more recent.
Eurasian backflow into Africa thus appears to have been a recurrent event in the history of many Africans, given its considerable impact on their genomes. Although population mixture in general is a process that increases genetic diversity, we observed a decrease in heterozygosity in the admixed Africans. Our simulations showed that these results are expected after mixture at these proportions with the Eurasians who suffered a significant bottleneck at the time of their exodus from Africa ∼60,000 ya. Consequently, we found that mixture can complicate interpretation of the coalescent history inferred from models that use the density of heterozygous sites in their implementations. In addition, we detected in admixed Africans an inflation of positive-selection signals on alleles associated with adult lactose tolerance and pigmentation in Europeans, but we suggest that these alleles have drifted neutrally in Africans after admixture. Furthermore, we detected Neanderthal ancestry in admixed Africans and found it to be proportional to their Eurasian ancestry. Similarly, in admixed Near Easterners, we found a decrease in Neanderthal ancestry proportional to the gene flow they have received from Africans. Although a higher genetic affinity of Neanderthals to Europeans than to Near Easterners was previously interpreted as additional Neanderthal admixture in the history of Europeans,42 we propose that a more parsimonious explanation for these observations is that African-Eurasian mixtures both introduced Neanderthal ancestry to Africa and “diluted” the Neanderthal ancestry in the Near East.
Am J Hum Genet. 2016 Dec 1; 99(6): 1316–1324.