

Post by Admin on Feb 24, 2023 21:18:55 GMT
Haplogroup R-M269 is the sub-clade of human Y-chromosome haplogroup R1b that is defined by the SNP marker M269. According to ISOGG 2020 it is phylogenetically classified as R1b1a1b. It underwent intensive research and was previously classified as R1b1a2 (2003 to 2005), R1b1c (2005 to 2008), R1b1b2 (2008 to 2011) and R1b1a1a2 (2011 to 2020).
According to a 2015 study, a hunter-gatherer from Samara (dated 5640-5555 cal BCE) belonging to haplogroup R1b1(*) was ancestral for both haplogroups R-M269 and R-M478. According to the authors, the occurrence of basal forms of R1b in eastern European hunter-gatherers provides a "geographically plausible source" for haplogroup R-M269. Subclades of R-M269, such as R-Z2103, have been found to be prevalent in ancient DNA found in individuals associated with the Yamnaya culture and related populations, and the dispersal of this haplogroup is associated with the spread of so-called "steppe ancestry" and at least some of the Indo-European languages.
A major Y-chromosome haplogroup R1b Holocene era founder effect in Central and Western Europe
Abstract
The phylogenetic relationships of numerous branches within the core Y-chromosome haplogroup R-M207 support a West Asian origin of haplogroup R1b, its initial differentiation there followed by a rapid spread of one of its sub-clades carrying the M269 mutation to Europe. Here, we present phylogeographically resolved data for 2043 M269-derived Y-chromosomes from 118 West Asian and European populations assessed for the M412 SNP that largely separates the majority of Central and West European R1b lineages from those observed in Eastern Europe, the Circum-Uralic region, the Near East, the Caucasus and Pakistan. Within the M412 dichotomy, the major S116 sub-clade shows a frequency peak in the upper Danube basin and Paris area with declining frequency toward Italy, Iberia, Southern France and British Isles. Although this frequency pattern closely approximates the spread of the Linearbandkeramik (LBK), Neolithic culture, an advent leading to a number of pre-historic cultural developments during the past ≤10 thousand years, more complex pre-Neolithic scenarios remain possible for the L23(xM412) components in Southeast Europe and elsewhere.
www.ncbi.nlm.nih.gov/pmc/articles/PMC3039512/
Genetic differentiation between upland and lowland populations shapes the Y-chromosomal landscape of West Asia
Abstract
Y-chromosomal variation in West Asian populations has so far been studied in less detail than in the neighboring Europe. Here, we analyzed 598 Y-chromosomes from two West Asian subregions—Transcaucasia and the Armenian plateau—using 40 Y-SNPs and 17 Y-STRs and combined them with previously published data from the region. The West Asian populations fell into two clusters: upland populations from the Anatolian, Armenian and Iranian plateaus, and lowland populations from the Levant, Mesopotamia and the Arabian Peninsula. This geographic subdivision corresponds with the linguistic difference between Indo-European and Turkic speakers, on the one hand, and Semitic speakers, on the other. This subdivision could be traced back to the Neolithic epoch, when upland populations from the Anatolian and Iranian plateaus carried similar haplogroup spectra but did not overlap with lowland populations from the Levant. We also found that the initial gene pool of the Armenian motherland population has been well preserved in most groups of the Armenian Diaspora. In view of the contribution of West Asians to the autosomal gene pool of the steppe Yamnaya archaeological culture, we sequenced a large portion of the Y-chromosome in haplogroup R1b samples from present-day East European steppe populations. The ancient Yamnaya samples are located on the “eastern” R-GG400 branch of haplogroup R1b-L23, showing that the paternal descendants of the Yamnaya still live in the Pontic steppe and that the ancient Yamnaya population was not an important source of paternal lineages in present-day West Europeans.
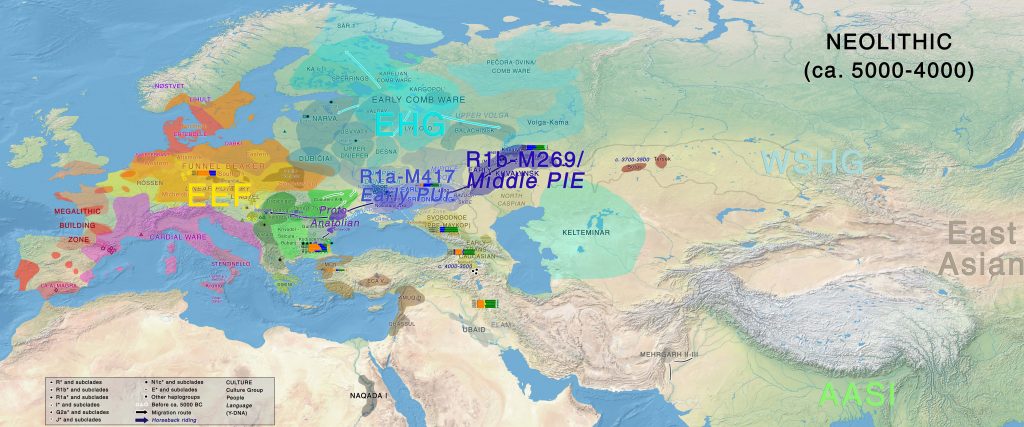
Neolithic
The development of a Middle PIE or Indo-Anatolian-speaking community must be identified with the peoples inhabiting the Volga-Ural region – i.e. probably Samara / early Khvalynsk cultures, whereas Early Proto-Uralic should probably be identified with cultures in the North Pontic region (such as early Sredni Stog), during the fifth millennium BC, in light of their cultural differences, their known samples and successive migration waves.
The different contribution of CHG ancestry from the Northern Caucasus, potentially from absorption of steppe peoples (apart from other potential admixture events) defines the formation of different Neolithic cultures, together with the appearance of evident Y-chromosome bottlenecks caused by expanding patrilineal clans and social interaction. The transition to nomadic pastoralism must have strengthened this social hierarchical trend that began ca. 5200-5000 BC in the steppe (Anthony 2007).
At the end of this period of initial differentiation (ca. 4500 BC or possibly earlier), the Suvorovo-Novodanilovka horse-rider chieftains appear in the North Pontic area and in the Lower Danube, bringing Khvalynsk symbology, which is likely to be the sign of migrating Proto-Anatolian speakers into the Balkans.
Eneolithic
Late Khvalynsk and early Repin cultural materials develop side by side in the Volga-Ural region representing the development of an early Late PIE community (Common Indo-European stage of Late PIE), while cultures to the west of the Don such as Dereivka or Kvitjana continue the early Sredni Stog traditions, thus probably connected with Proto-Uralic on the forest-steppe region. The Caucasus Mountains remains an ethnolinguistic frontier between the steppe and North Iran, although some influence is seen from the steppe into Maykop, from known economic contacts.
Migration waves from late Repin / Early Yamna from ca. 3500/3400 BC are seen initially on the Northern Caucasus and the Lower Volga – Ural regions, and eventually in different samples to the east in Central Asia, and to the south in Iran. Almost all of them share R1b-L23 lineages. Pre-Tocharian (of a more archaic Late PIE nature) is probably to be identified with these migration waves into the Altai region, developing the Afanasevo culture (predominantly of R1b-L23 lineages).
At the same time, late Repin migrants settle as clear “colonizers” to the west of the Don River (displacing cultures from the North Pontic region), to form the western South-Bug / Lower Don groups, which – together with the Volga-Ural / North Caucasian groups – become part the early Yamna culture, which dominates from ca. 3300 BC over the Pontic-Caspian steppe. To the north of the North Pontic steppe-forest or forest zone, a Uralic-speaking Proto-Corded Ware culture develops (probably related to displaced North Pontic forest-steppe pastoralists) somewhere in or around the Dnieper-Dniester-Bug-Vistula area of interaction with late Trypillia, TRB, and GAC cultures.
According to a 2015 study, a hunter-gatherer from Samara (dated 5640-5555 cal BCE) belonging to haplogroup R1b1(*) was ancestral for both haplogroups R-M269 and R-M478. According to the authors, the occurrence of basal forms of R1b in eastern European hunter-gatherers provides a "geographically plausible source" for haplogroup R-M269. Subclades of R-M269, such as R-Z2103, have been found to be prevalent in ancient DNA found in individuals associated with the Yamnaya culture and related populations, and the dispersal of this haplogroup is associated with the spread of so-called "steppe ancestry" and at least some of the Indo-European languages.
A major Y-chromosome haplogroup R1b Holocene era founder effect in Central and Western Europe
Abstract
The phylogenetic relationships of numerous branches within the core Y-chromosome haplogroup R-M207 support a West Asian origin of haplogroup R1b, its initial differentiation there followed by a rapid spread of one of its sub-clades carrying the M269 mutation to Europe. Here, we present phylogeographically resolved data for 2043 M269-derived Y-chromosomes from 118 West Asian and European populations assessed for the M412 SNP that largely separates the majority of Central and West European R1b lineages from those observed in Eastern Europe, the Circum-Uralic region, the Near East, the Caucasus and Pakistan. Within the M412 dichotomy, the major S116 sub-clade shows a frequency peak in the upper Danube basin and Paris area with declining frequency toward Italy, Iberia, Southern France and British Isles. Although this frequency pattern closely approximates the spread of the Linearbandkeramik (LBK), Neolithic culture, an advent leading to a number of pre-historic cultural developments during the past ≤10 thousand years, more complex pre-Neolithic scenarios remain possible for the L23(xM412) components in Southeast Europe and elsewhere.
www.ncbi.nlm.nih.gov/pmc/articles/PMC3039512/
Genetic differentiation between upland and lowland populations shapes the Y-chromosomal landscape of West Asia
Abstract
Y-chromosomal variation in West Asian populations has so far been studied in less detail than in the neighboring Europe. Here, we analyzed 598 Y-chromosomes from two West Asian subregions—Transcaucasia and the Armenian plateau—using 40 Y-SNPs and 17 Y-STRs and combined them with previously published data from the region. The West Asian populations fell into two clusters: upland populations from the Anatolian, Armenian and Iranian plateaus, and lowland populations from the Levant, Mesopotamia and the Arabian Peninsula. This geographic subdivision corresponds with the linguistic difference between Indo-European and Turkic speakers, on the one hand, and Semitic speakers, on the other. This subdivision could be traced back to the Neolithic epoch, when upland populations from the Anatolian and Iranian plateaus carried similar haplogroup spectra but did not overlap with lowland populations from the Levant. We also found that the initial gene pool of the Armenian motherland population has been well preserved in most groups of the Armenian Diaspora. In view of the contribution of West Asians to the autosomal gene pool of the steppe Yamnaya archaeological culture, we sequenced a large portion of the Y-chromosome in haplogroup R1b samples from present-day East European steppe populations. The ancient Yamnaya samples are located on the “eastern” R-GG400 branch of haplogroup R1b-L23, showing that the paternal descendants of the Yamnaya still live in the Pontic steppe and that the ancient Yamnaya population was not an important source of paternal lineages in present-day West Europeans.
Neolithic
The development of a Middle PIE or Indo-Anatolian-speaking community must be identified with the peoples inhabiting the Volga-Ural region – i.e. probably Samara / early Khvalynsk cultures, whereas Early Proto-Uralic should probably be identified with cultures in the North Pontic region (such as early Sredni Stog), during the fifth millennium BC, in light of their cultural differences, their known samples and successive migration waves.
The different contribution of CHG ancestry from the Northern Caucasus, potentially from absorption of steppe peoples (apart from other potential admixture events) defines the formation of different Neolithic cultures, together with the appearance of evident Y-chromosome bottlenecks caused by expanding patrilineal clans and social interaction. The transition to nomadic pastoralism must have strengthened this social hierarchical trend that began ca. 5200-5000 BC in the steppe (Anthony 2007).
At the end of this period of initial differentiation (ca. 4500 BC or possibly earlier), the Suvorovo-Novodanilovka horse-rider chieftains appear in the North Pontic area and in the Lower Danube, bringing Khvalynsk symbology, which is likely to be the sign of migrating Proto-Anatolian speakers into the Balkans.
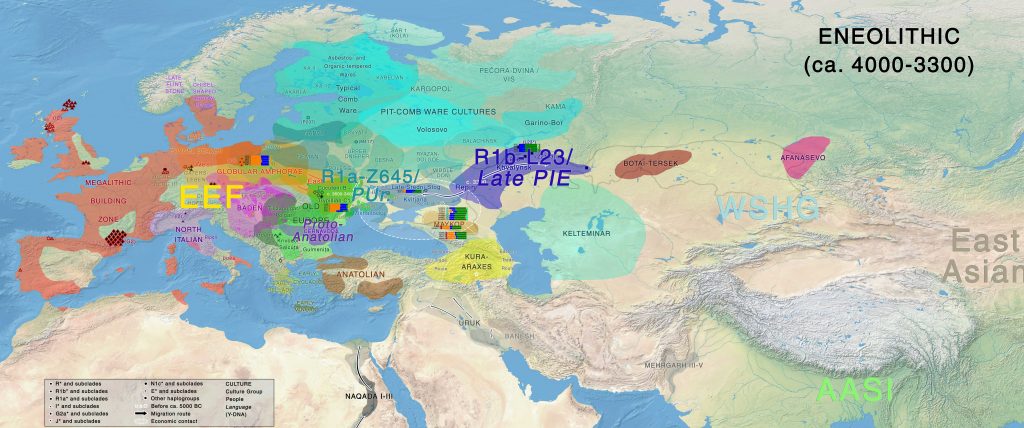
Eneolithic
Late Khvalynsk and early Repin cultural materials develop side by side in the Volga-Ural region representing the development of an early Late PIE community (Common Indo-European stage of Late PIE), while cultures to the west of the Don such as Dereivka or Kvitjana continue the early Sredni Stog traditions, thus probably connected with Proto-Uralic on the forest-steppe region. The Caucasus Mountains remains an ethnolinguistic frontier between the steppe and North Iran, although some influence is seen from the steppe into Maykop, from known economic contacts.
Migration waves from late Repin / Early Yamna from ca. 3500/3400 BC are seen initially on the Northern Caucasus and the Lower Volga – Ural regions, and eventually in different samples to the east in Central Asia, and to the south in Iran. Almost all of them share R1b-L23 lineages. Pre-Tocharian (of a more archaic Late PIE nature) is probably to be identified with these migration waves into the Altai region, developing the Afanasevo culture (predominantly of R1b-L23 lineages).
At the same time, late Repin migrants settle as clear “colonizers” to the west of the Don River (displacing cultures from the North Pontic region), to form the western South-Bug / Lower Don groups, which – together with the Volga-Ural / North Caucasian groups – become part the early Yamna culture, which dominates from ca. 3300 BC over the Pontic-Caspian steppe. To the north of the North Pontic steppe-forest or forest zone, a Uralic-speaking Proto-Corded Ware culture develops (probably related to displaced North Pontic forest-steppe pastoralists) somewhere in or around the Dnieper-Dniester-Bug-Vistula area of interaction with late Trypillia, TRB, and GAC cultures.


