|
|
Post by Admin on Jan 28, 2019 23:44:09 GMT
Over the past 30 years, understanding of Homo sapiens evolution has advanced greatly. Most research has supported the theory that modern humans had originated in Africa by about 200,000 years ago, but the latest findings reveal more complexity than anticipated. They confirm interbreeding between H. sapiens and other hominin species, provide evidence for H. sapiens in Morocco as early as 300,000 years ago, and reveal a seemingly incremental evolution of H. sapiens cranial shape. Although the cumulative evidence still suggests that all modern humans are descended from African H. sapiens populations that replaced local populations of archaic humans, models of modern human origins must now include substantial interactions with those populations before they went extinct. These recent findings illustrate why researchers must remain open to challenging the prevailing theories of modern human origins. At an archaeological site near the Atlantic coast, finds of skull, face and jaw bones identified as being from early members of our species have been dated to about 315,000 years ago. That indicates H. sapiens appeared more than 100,000 years earlier than thought: most researchers have placed the origins of our species in East Africa about 200,000 years ago. “Until now, the common wisdom was that our species emerged probably rather quickly somewhere in a ‘Garden of Eden’ that was located most likely in sub-Saharan Africa,” says Jean-Jacques Hublin, an author of the study and a director at the Max Planck Institute for Evolutionary Anthropology in Leipzig, Germany. Now, “I would say the Garden of Eden in Africa is probably Africa — and it’s a big, big garden.” Hublin was one of the leaders of the decade-long excavation at the Moroccan site, called Jebel Irhoud. 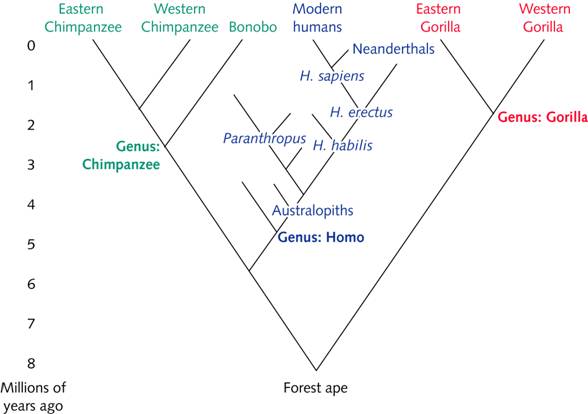 The re-dating and the tranche of new human bones convince Hublin that early H. sapiens once lived at Jebel Irhoud. “It’s a face you could cross in the street today,” he says. The teeth — although big compared with those of today's humans — are a better match to H. sapiens than they are to Neanderthals or other archaic humans. And the Jebel Irhoud skulls, elongated compared with those of later H. sapiens, suggest that these individuals' brains were organized differently. This offers clues about the evolution of the H. sapiens lineage into today’s anatomically modern humans. Hublin suggests that anatomically modern humans may have acquired their characteristic faces before changes to the shape of their brains occurred. Moreover, the mix of features seen in the Jebel Irhoud remains and other H. sapiens-like fossils from elsewhere in Africa point to a diverse genesis for our species, and raises doubt about an exclusively East African origin. “Homo sapiens, despite being so well known, was a species without a past until now,” says María Martínon-Torres, a palaeoanthropologist at University College London, noting the scarcity of fossils linked to human origins in Africa. But the lack of features that, she says, define our species — such as a prominent chin and forehead — convince her that the Jebel Irhoud remains should not be considered H. sapiens. Chris Stringer, a palaeoanthropologist at the Natural History Museum in London, who co-authored a News & Views article accompanying the studies, says he was baffled by the Jebel Irhoud remains when he first saw them in the early 1970s. He knew that they weren’t Neanderthals, but they seemed too young and primitive-looking to be H. sapiens. But with the older dates and the new bones, Stringer agrees that the Jebel Irhoud bones stand firmly on the H. sapiens lineage. “They shift Morocco from a supposed backwater in the evolution of our species to a prominent position,” he adds. |
|
|
|
Post by Admin on Jan 29, 2019 18:48:17 GMT
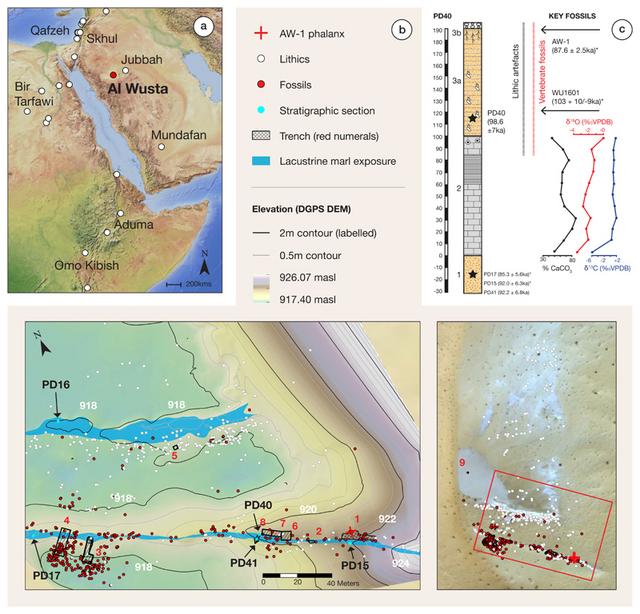 Figure 1 Al Wusta location, map of site and stratigraphy. Homo sapiens evolved in Africa in the late Middle Pleistocene1. Early dispersals out of Africa are evidenced at the Levantine site of Misliya at ~194-177 ka2, followed by Skhul and Qafzeh, where H. sapiens fossils have been dated to ~130-100 and ~100-90 ka respectively3. While the Levantine fossil evidence has been viewed as the onset of a much broader dispersal into Asia4–6, it has generally been seen as representing short-lived incursions into the woodlands of the Levant immediately adjacent to Africa, where relatively high precipitation is produced by winter storms tracking across the Mediterranean7,8. While the Levantine record indicates the subsequent local replacement of early H. sapiens by Neanderthals, the failure of early dispersals to extend beyond the Levant is largely inferred from interpretations of genetic data9. Genetic studies have suggested that recent non-African populations stem largely10, if not entirely9, from an expansion ~60-50 ka, but this model remains debated. The absence of low latitude Pleistocene human DNA and uncertainties regarding ancient population structure undermine conclusions drawn from genetic studies alone. The paucity of securely dated archaeological, palaeontological and ancient DNA data - particularly across southern Asia - has made testing dispersal hypotheses challenging4,7,11. The Arabian Peninsula is a vast landmass at the crossroads of Africa and Eurasia. Growing archaeological evidence demonstrates repeated hominin occupations of Arabia20,21 each associated with a strengthened summer monsoon which led to the re-activation of lakes and rivers22–24, as it did in North Africa25. Here we report the discovery of the first pre-Holocene human fossil in Arabia, Al Wusta-1 (AW-1), as well as the age, stratigraphy, vertebrate fossils and stone tools at the Al Wusta site (Fig. 1, see also Supplementary Information). 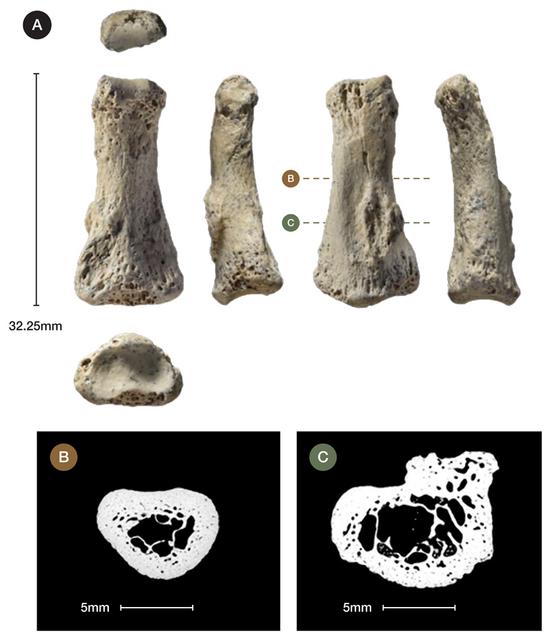 Figure 2 Photographs and micro-CT scans of Al Wusta-1 Homo sapiens phalanx. AW-1 is an intermediate manual phalanx, most likely from the 3rd ray (Fig. 2a, Supplementary Information 1: see below for detail on siding and species identification). It is generally well-preserved, although there is some erosion of the cortical/subchondral bone, and minor pathological bone formation (likely an enthesophyte) affecting part of the diaphysis (Supplementary Information 1). The phalanx measures 32.3 mm in proximo-distal length, and 8.7 mm and 8.5mm in radio-ulnar breadth of the proximal base and midshaft, respectively (Supplementary Table 1). AW-1 is more gracile than the robust intermediate phalanges of Neanderthals26–28, which are broader radio-ulnarly relative to their length and have a more 'flared' base. AW-1’s proximal radio-ulnar maximum breadth is 14.98 mm, which provides an intermediate phalanx breadth-length index (proximal radio-ulnar maximum breadth relative to articular length) of 49.6. This is very similar to the mean (± SD) for the Skhul and Qafzeh H. sapiens of 49.7 (± 4.1) and 49.1 (± 4.0) for Upper Palaeolithic Europeans, but 1.89 standard deviations below the Neanderthal mean of 58.3 (± 4.6)29. To provide a broad interpretive context for the Al Wusta phalanx, we conducted linear and geometric morphometric (GMM) landmark analyses (Supplementary Information 1) on phalanges from non-human primates, fossil hominins and geographically widespread recent H. sapiens. Comparative linear analyses (Supplementary Information 1, Supplementary Tables 2 and 3, Supplementary Figure 1) reveal that there is substantial overlap across most taxa for all shape ratios, so AW-1 falls within the range of variation of H. sapiens, cercopiths, Gorilla, Australipithecus afarensis, A. sediba and Neanderthals. However, AW-1 is most similar to the median value or falls within the range of variation of recent and early H. sapiens for all shape ratios. |
|
|
|
Post by Admin on Jan 30, 2019 18:16:23 GMT
Geometric morphometric (GMM) analyses of AW-1 and various primate groups including hominins (see Supplementary Table 4 and Supplementary Figure 2 for landmarks, and Supplementary Table 5 for sample) are illustrated in Figure 3 and Supplementary Figure 3. PC1 and PC2 together account for 61% of group variance in shape. AW-1 is separated on these two shape vectors from the non-human primates and most of the Neanderthals. AW-1 falls closest to the recent and early H. sapiens and is clearly differentiated from all non-human primates. This is also shown by the Procrustes distances from AW-1 to the mean shapes of each taxonomic group (Supplementary Table 6). 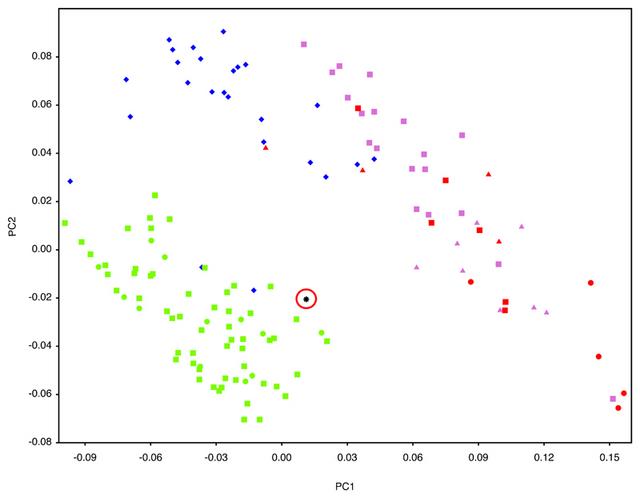 Figure 3 Scatterplot of the first two principal components (PC) scores of the geometric morphometric analysis of the Al Wusta-1 phalanx compared with a sample of primates, including hominins. Three of the Neanderthal phalanges (from Kebara 2 and Tabun C1) are quite disparate from the main Neanderthal cluster and fall closer to the H. sapiens and Al Wusta cluster on PC1 and 2 (Figure 3 and Supplementary Figure 3). Having established the hominin affinity of AW-1, shape was analysed in more detail using a smaller hominin sample for which ray number and side were known, which included Kebara 2 and Tabun C1. The broader primate sample used in the first GMM analysis was not used for the more detailed shape analysis, as the initial comparisons show clearly that AW-1 is not a non-human primate and including this level of variation could potentially mask more subtle shape differences between hominins. The side and ray are also not known for most of the Neanderthal and non-human primate samples, meaning it would be impossible to evaluate the effect of these factors using this sample. The more in-depth shape comparison and modelling using the hominin sample of phalanges of known ray and side (Supplementary Table 7) demonstrates that the long and slender morphology of AW-1 falls just outside the range of variation of comparative Middle Palaeolithic modern humans, but that its affinity is clearly with H. sapiens rather than Neanderthals (Fig. 4, Supplementary Table 8). Although both Pleistocene H. sapiens and Neanderthal landmark configurations fall almost completely inside the scatter for the Holocene H. sapiens sample in the principal components analysis (Figure 4), AW-1 is closest to Holocene H. sapiens 3rd intermediate phalanges. AW-1 overlaps with the Holocene H. sapiens sample, but is separated from the Pleistocene H. sapiens specimens by a higher score on PC2 and from the Neanderthal group by a simultaneously higher score on PC1 and PC2.  This Neanderthal woman was found in Gibraltar. The Neanderthals are the closest extinct human relative. They went extinct about 40,000 years ago. The Procrustes distances (Supplementary Table 8), also show that AW-1 is most distinct from the Neanderthal phalanges, which fall towards the lower ends of both PCs and are characterised by shorter and broader dimensions. PC1 and PC2 in this analysis show that AW-1 is taller and narrower (in all directions: dorso-palmarly, proximo-distally and radio-ulnarly) than almost all the phalanges in the comparative sample and is particularly distinct from most of the Neanderthal phalanges. In this analysis AW-1 is closest in shape to 3rd phalanges of individuals from (in descending order of proximity) Egyptian Nubia, and Medieval Canterbury (UK), and Maiden Castle (Iron Age Dorset, UK) (Supplementary Table 9), although there is not a great difference in its distance to any of these specimens. These analyses suggest that the AW-1 phalanx is likely to be a 3rd intermediate phalanx from a H. sapiens individual. |
|
|
|
Post by Admin on Jan 31, 2019 19:34:42 GMT
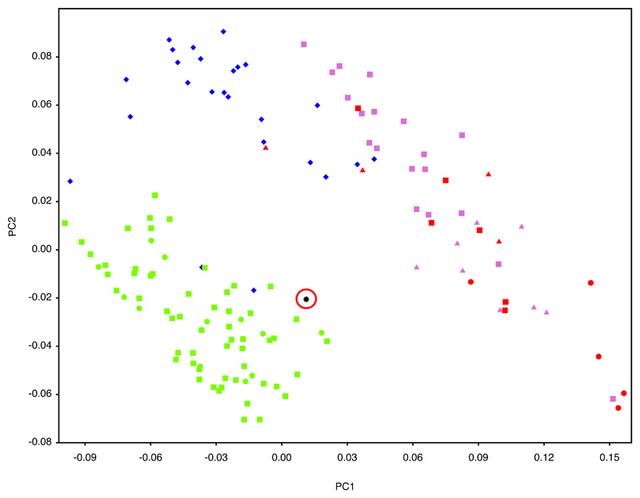 Figure 4 Scatterplot of the first two principal component (PC) scores from the geometric morphometric analyses of AW-1 and sample of comparative hominin 2nd, 3rd, and 4th intermediate phalanges. The more in-depth shape comparison and modelling using the hominin sample of phalanges of known ray and side (Supplementary Table 7) demonstrates that the long and slender morphology of AW-1 falls just outside the range of variation of comparative Middle Palaeolithic modern humans, but that its affinity is clearly with H. sapiens rather than Neanderthals (Fig. 4, Supplementary Table 8). Although both Pleistocene H. sapiens and Neanderthal landmark configurations fall almost completely inside the scatter for the Holocene H. sapiens sample in the principal components analysis (Figure 4), AW-1 is closest to Holocene H. sapiens 3rd intermediate phalanges. AW-1 overlaps with the Holocene H. sapiens sample, but is separated from the Pleistocene H. sapiens specimens by a higher score on PC2 and from the Neanderthal group by a simultaneously higher score on PC1 and PC2. The Procrustes distances (Supplementary Table 8), also show that AW-1 is most distinct from the Neanderthal phalanges, which fall towards the lower ends of both PCs and are characterised by shorter and broader dimensions. PC1 and PC2 in this analysis show that AW-1 is taller and narrower (in all directions: dorso-palmarly, proximo-distally and radio-ulnarly) than almost all the phalanges in the comparative sample and is particularly distinct from most of the Neanderthal phalanges. In this analysis AW-1 is closest in shape to 3rd phalanges of individuals from (in descending order of proximity) Egyptian Nubia, and Medieval Canterbury (UK), and Maiden Castle (Iron Age Dorset, UK) (Supplementary Table 9), although there is not a great difference in its distance to any of these specimens. These analyses suggest that the AW-1 phalanx is likely to be a 3rd intermediate phalanx from a H. sapiens individual. The third ray is the most symmetrical ray in the hand and is therefore difficult to side, particularly when not all of the phalanges of a particular individual are present. Comparing AW-1 separately to right and to left phalanges (Supplementary Information 1.4) gives results which are very similar to the pooled sample, such that AW-1 is closest to Holocene H. sapiens 3rd rays for both right and left hands (Supplementary Figure 4, Supplementary Table 10). There is little difference in morphological closeness between AW-1 and its nearest neighbour in the samples of right and left bones (Supplementary Table 11), reflecting the lack of difference in morphology between the sides. It is therefore not possible to suggest whether AW-1 comes from a right or a left hand using these analyses. AW-1 is unusual in its more circular midshaft cross-sectional shape (Fig. 2B), which is confirmed by cross-sectional geometric analyses (Supplementary Information 1.5). This may reflect the pronounced palmar median bar that makes the palmar surface slightly convex at the midshaft rather than flat, the latter being typical of most later Homo intermediate phalanges. However, more circular shafts may reflect greater loading of the bone in multiple directions and enthesophytes are a common response to stress from high levels of physical activity30. This morphology may reflect high and varied loading of the fingers during intense manual activity. To determine the age of AW-1, and associated sediments and fossils, we used a combination of uranium series (U-series), electron spin resonance (ESR) and optically stimulated luminescence (OSL) dating (Methods, Supplementary Information 2 and 3). U-series ages were produced for AW-1 itself (87.6 ± 2.5 ka) and hippopotamus dental tissues (WU1601), which yielded ages of 83.5 ± 8.1 ka (enamel) and 65.0 ± 2.1 ka (dentine). They should be regarded as minimum estimates for the age of the fossils. In addition, a combined U-series-ESR age calculation for WU1601 yielded an age of 103 +10/-9 ka. AW-1 was found on an exposure of Unit 3b, and WU1601 excavated from Unit 3a, one metre away (Fig 1b). Unit 1 yielded OSL ages of 85.3 ± 5.6 ka (PD17), 92.2 ± 6.8 ka (PD41) and 92.0 ± 6.3 ka (PD15), while Unit 3a yielded an OSL age of 98.6 ± 7.0 ka (PD40). The OSL age estimates agree within error with the US-ESR age obtained for WU-1601 and the minimum age of ~88 ka obtained for AW-1. These data were incorporated into a Bayesian sequential phase model31 which indicates that deposition of Unit 1 ceased 93.1 ± 2.6 ka (Phase 1: PD15, 17, 41) and that Units 2 and 3 and all associated fossils were deposited between 92.2 ± 2.6 ka and 90.4 ± 3.9 ka (Phase 2: all other ages) (Supplementary Information 4, Supplementary Figure 11).  Figure 5 Selected Al Wusta lithic artefacts. This ~95-86 ka timeframe is slightly earlier than most other records of increased humidity in the region in late MIS 5 32,33, which correlate with a strengthened summer monsoon associated with an insolation peak at 84 ka (Fig. 6). The underlying (Unit 3) aeolian sand layer at Al Wusta correlates with an insolation minimum at the end of MIS 5c. The chronometric age estimates for the site suggest that lake formation and the associated fauna and human occupation occurred shortly after this in time. Regional indications of increased humidity around the 84 ka insolation peak include speleothem formation at ~88 ka in the Negev34, and the formation of sapropel S3 beginning ~86 ka35. In both the Levant and Arabia, records are consistent with this switch from aridity to humidity around this time32–40. Precisely reconstructing regional palaeoclimate at this time and relating it to human demographic and behavioural change has proved challenging. This reflects both rapid changes in climate, as well as the complexities involved in dating relevant deposits41. In summary, combining chronological data (Supplementary sections 2-4), interpretation of the sedimentary sequence (described below), and the regional setting of Al Wusta, we conclude that lake formation and associated finds such as the AW-1 phalanx relate to the late MIS 5 humid period associated with the 84 ka insolation peak. 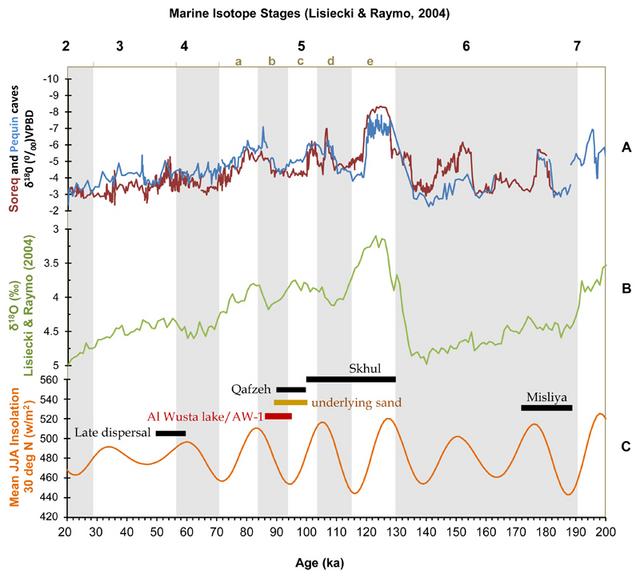 Figure 6 The chronological and climatic context of Al Wusta. Al Wusta-1 is the oldest directly dated H. sapiens fossil outside Africa and the Levant. It joins a small but growing corpus of evidence that the early dispersal of H. sapiens into Eurasia was much more widespread than previously thought. The site of Al Wusta is located in the Nefud desert more than 650 km southeast of Skhul and Qafzeh (Fig. 1A). This site establishes that H. sapiens were in Arabia in late MIS 5, rather than being restricted to Africa and the Levant as suggested by traditional models (Fig. 6). With Skhul dating to ~130-100 ka, Qafzeh to ~100-90 ka3,46 and Al Wusta to ~95-85 ka it is currently unclear if the southwest Asian record reflects multiple early dispersals out of Africa or a long occupation during MIS 5. The association of the Al Wusta site with a late MIS 5 humid phase (Fig. 6), suggests that significant aspects of this dispersal process were facilitated by enhanced monsoonal rainfall. While changes in behaviour and demography are crucial to understanding the dispersal process, climatic windows of opportunity were also key in allowing H. sapiens to cross the Saharo-Arabian arid belt, which often constituted a formidable barrier24,25. Al Wusta shows that the early, Marine Isotope Stage 5, dispersals of H. sapiens out of Africa were not limited to the Levantine woodlands sustained by winter rainfall, but extended deep into the Arabian interior where enhanced summer rainfall created semi-arid grasslands containing abundant fauna and perennial lakes. After long being isolated in Africa1,47,48, the Late Pleistocene saw the expansion of our species out of Africa and into the diverse ecologies of Eurasia. Within a few thousand years of spreading into Eurasia our species was occupying rainforest environments and making long sea crossings to remote islands13,18. Adapting to the semi-arid conditions of the Saharo-Arabian arid belt represented a crucial step on this pathway to global success and the Al Wusta Homo sapiens fossil demonstrates this early ability to occupy diverse ecologies which led to us becoming a cosmopolitan species. Nat Ecol Evol. 2018 May; 2(5): 800–809. |
|
|
|
Post by Admin on Feb 8, 2019 17:23:51 GMT
:focal(1000x800:1001x801)/https://public-media.si-cdn.com/filer/59/e2/59e216c7-6f22-4969-b408-063d0e73821d/ho_skull_casts_front.jpg) The study, published in Nature Communications, is one of the first examples of how machine learning can help reveal clues to our own origins. By poring through vast amounts of genomic data left behind in fossilized bones and comparing it with DNA in modern humans, scientists can begin to fill in some of the many gaps of our species’ evolutionary history. In this case, the results seem to match paleoanthropology theories that were developed from studying human ancestor fossils found in the ground. The new data suggest that the mysterious hominin was likely descended from an admixture of Neanderthals and Denisovans (who were only identified as a unique species on the human family tree in 2010). Such a species in our evolutionary past would look a lot like the fossil of a 90,000-year-old teenage girl from Siberia's Denisova cave. Her remains were described last summer as the only known example of a first-generation hybrid between the two species, with a Neanderthal mother and a Denisovan father.  Previous human genome studies have revealed that after modern humans left Africa, perhaps 180,000 years ago, they subsequently interbred with species like Neanderthals and Denisovans, who coexisted with early modern humans before going extinct. But redrawing our family tree to include these divergent branches has been difficult. Evidence for “ghost” species can be sparse, and many competing theories exist to explain when, where, and how often Homo sapiens might have interbred with other species. Traces of these ancient interspecies liaisons, called introgressions, can be identified as places of divergence in the human genome. Scientists observe more separation between two chromosomes than you'd expect if both of the chromosomes came from the same human species. When scientists sequenced the Neanderthal genome in 2010, they realized that some of these divergences represented fractions of our genome that came from Neanderthals. Studies have also revealed that some living humans can trace as much as 5 percent of their ancestry to Denisovans.  Identifying and analyzing the many divergent places throughout the genome, and computing the countless genetic combinations that could have produced them, is too big a job for humans to tackle on their own—but it's a task that may be tailor made for deep learning algorithms. Deep learning is a type of artificial intelligence in which algorithms are designed to work as an artificial neural network, or a program that can process information the same way a mammalian brain would. These machine learning systems can detect patterns and account for previous information to “learn,” allowing them to perform new tasks or look for new information after analyzing enormous amounts of data. (A common example is Google DeepMind’s AlphaZero, which can teach itself to master board games.) 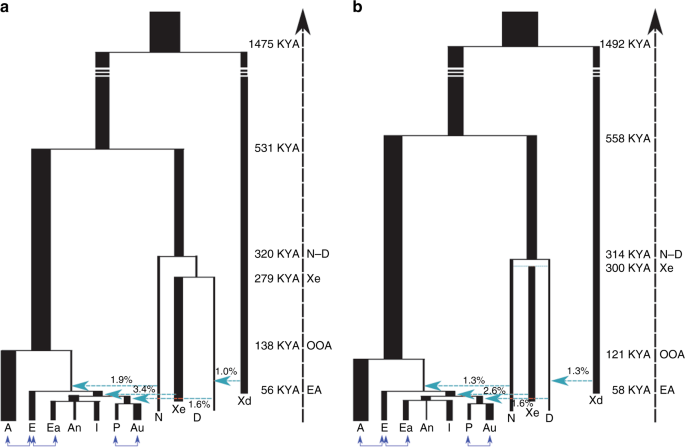 In this case, machines were set to work analyzing the human genome and predicting human demographics by simulating how our DNA might have evolved over many thousands of possible scenarios of ancient evolution. The program accounted for the structure and evolution of DNA as well as models of human migration and interbreeding to try to fit some of the pieces together in an incredibly complex puzzle. The researchers trained the computer to analyze eight different models of the most plausible theories of early human evolution across Eurasia. The models came from previous studies that attempted to come up with a scenario that would result in the current picture of the human genome, including its known Neanderthal and Denisovan components. |
|

