|
|
Post by Admin on Mar 4, 2021 19:37:52 GMT
Preliminary paleohistological observations of the StW 573 (‘Little Foot’) skull
Abstract
Numerous aspects of early hominin biology remain debated or simply unknown. However, recent developments in high-resolution imaging techniques have opened new avenues in the field of paleoanthropology. More specifically, X-ray synchrotron-based analytical imaging techniques have the potential to provide crucial details on the ontogeny, physiology, biomechanics, and biological identity of fossil specimens. Here we present preliminary results of our X-ray synchrotron-based investigation of the skull of the 3.67-million-year-old Australopithecus specimen StW 573 (‘Little Foot’) at the I12 beamline of the Diamond Light Source (United Kingdom). Besides showing fine details of the enamel (i.e., hypoplasias) and cementum (i.e., incremental lines), as well as of the cranial bone microarchitecture (e.g., diploic channels), our synchrotron-based investigation reveals for the first time the 3D spatial organization of the Haversian systems in the mandibular symphysis of an early hominin.
Applications of X-ray synchrotron-based analytical techniques in evolutionary studies have opened up new avenues in the field of paleoanthropology. In particular, X-ray synchrotron microtomography has been proved to be particularly useful for imaging anatomical structures in extant and fossil hominins that are traditionally observed through destructive histological methods (e.g., Tafforeau and Smith, 2008; Maggiano et al., 2016; Andronowski et al., 2017; Mani-Caplazi et al., 2017; Gunz et al., 2020). For instance, microscopic analyses of fossil craniodental specimens using synchrotron radiation have revealed previously unknown aspects of the ontogeny of extinct hominin taxa (Tafforeau and Smith, 2008; Gunz et al., 2020).
Besides its geological age of 3.67 million years, StW 573 (‘Little Foot’) is remarkable for its outstanding degree of preservation and completeness (Clarke and Kuman, 2019). The high-resolution virtual exploration of the ‘Little Foot’ skull is thus expected to provide new insights into the Pliocene Australopithecus’ biology. Here, we present preliminary results of our X-ray synchrotron-based investigation of the dentition and cranial bones of ‘Little Foot’. The main aim of our study is to identify and assess the degree of preservation of craniodental microstructures that could contribute to the reconstruction of Australopithecus’ biology. To the best of our knowledge, this is the first time that histological features of the compact bone of a Pliocene hominin skull have been non-invasively observed.
|
|
|
|
Post by Admin on Mar 5, 2021 20:00:14 GMT
Introduction Applications of X-ray synchrotron-based analytical techniques in evolutionary studies have opened up new avenues in the field of paleoanthropology. In particular, X-ray synchrotron microtomography has been proved to be particularly useful for imaging anatomical structures in extant and fossil hominins that are traditionally observed through destructive histological methods (e.g., Tafforeau and Smith, 2008; Maggiano et al., 2016; Andronowski et al., 2017; Mani-Caplazi et al., 2017; Gunz et al., 2020). For instance, microscopic analyses of fossil craniodental specimens using synchrotron radiation have revealed previously unknown aspects of the ontogeny of extinct hominin taxa (Tafforeau and Smith, 2008; Gunz et al., 2020). Besides its geological age of 3.67 million years, StW 573 (‘Little Foot’) is remarkable for its outstanding degree of preservation and completeness (Clarke and Kuman, 2019). The high-resolution virtual exploration of the ‘Little Foot’ skull is thus expected to provide new insights into the Pliocene Australopithecus’ biology. Here, we present preliminary results of our X-ray synchrotron-based investigation of the dentition and cranial bones of ‘Little Foot’. The main aim of our study is to identify and assess the degree of preservation of craniodental microstructures that could contribute to the reconstruction of Australopithecus’ biology. To the best of our knowledge, this is the first time that histological features of the compact bone of a Pliocene hominin skull have been non-invasively observed. Histology of dental tissues Figure 1 shows two dimensional (2D) sections through the roots of the upper right first molar with a resolution of 3.25 µm. These sections reveal the presence of cementum between the dentine and sediments filling the tooth alveolus (Figure 1A,B). The dentine–cementum junction and the boundary between the cementum and the sediments are clearly visible. Cementum microstructures, such as incremental lines, are discernible (Figure 1B). Additionally, we reconstructed in three dimensions (3D) the enamel cap of the lower left canine using image stacks at 21.23 and 7.91 µm resolution (Figure 2). Lines and pits observable on the distal-buccal aspect of the 21.23-µm reconstruction are located at about 2.4 and 3.6 mm from the cemento-enamel junction, which represents 36% and 54% of the crown height, respectively. While fine details of the pits are clearly visible in the 3D reconstruction using synchrotron-based data sets at 7.91 µm, the color map reveals that they are slightly less clearly rendered in the 21.23-µm 3D reconstruction, especially on the mesial-buccal aspect (Figure 2C,D). These reductions in the normal thickness of enamel correspond to disruptions to the normal growth of enamel (i.e., hypoplasias) and indicate two disruptive events in StW 573’s life history. Figure 1 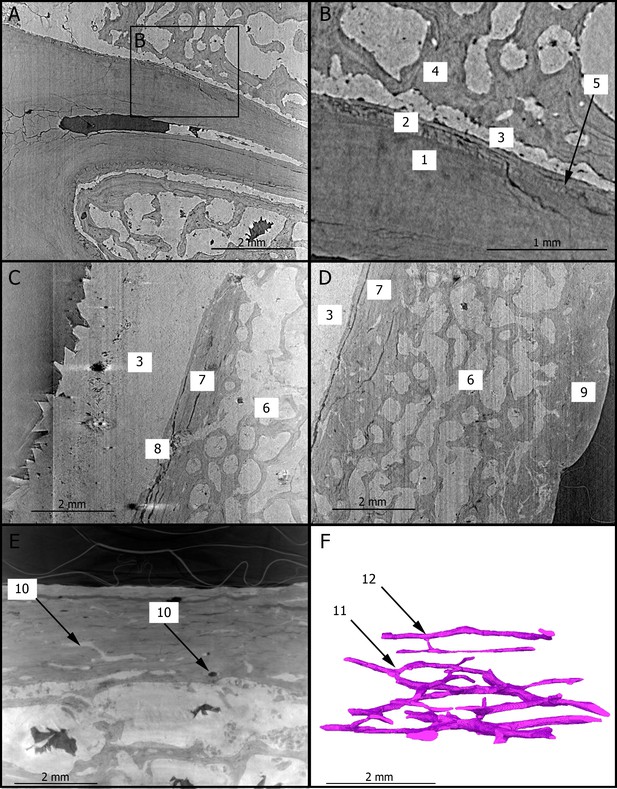 2D sections of the roots of the upper right first molar (A,B), of the cranial vault (C,D), and of the mandibular symphysis (E) of StW 573. Figure 2 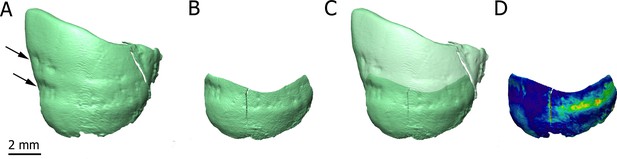 3D renderings of the buccal aspect of the enamel surface of the lower left canine of StW 573 using synchrotron-based data sets at 21.23 (A) and 7.91 µm (B). Histology of bone tissues Figure 1C–E show 2D sections through the cranial vault and the mandibular symphysis with a resolution of 3.25 µm. The voids in the spongious bone have been partially or completely filled by calcite. The opening of the diploic channel through the inner table in the cranial vault could be identified (Figure 1C). The Haversian system is discernible in the outer and inner tables of the cranial vault (Figure 1C,D) and clearly visible in the compact bone of the mandible (Figure 1E). Figure 1F shows a 3D reconstruction of the canal network and branching and interconnections between the Haversian systems. Based on the terminology defined by Maggiano et al., 2016, we identify a Type 2 (i.e., dichotomous) branching pattern. Furthermore, a linear transverse connection (i.e., Volkmann’s canal) can be observed. Vascular canals are proportionally more abundant close to the trabecular bone than in the rest of the compact bone (Figure 1F). The Haversian canals globally lie parallel to the external surface. |
|
|
|
Post by Admin on Mar 5, 2021 23:29:29 GMT
Collectively, our results show that the virtual histological investigation of both dental and bone tissues in complete fossil hominin skulls by using high-resolution synchrotron radiation may be possible. Contrary to traditional histological analyses based on physical sections of the bone (e.g., Bartsiokas, 2002) and imaging-based exploration of volumes of interest physically extracted from bones (e.g., Maggiano et al., 2016; Andronowski et al., 2017), this method, as previously demonstrated in landmark papers for dental microstructures using dentognathic remains (e.g., Tafforeau and Smith, 2008), offers the possibility of non-invasively investigating microscopic structures in complete crania that have deep implications for the ontogeny, physiology, biomechanics, and biological identity of fossil specimens.
More specifically, our observations reveal that ‘Little Foot’ preserves fine cementum microstructures that can be quantitatively explored. Because cementum is rarely remodeled during life, it may preserve valuable information about hominin paleobiology (rev. in Tang et al., 2016). In particular, the incremental lines may be used to determine age-at-death as well as specific stress periods that might be related to life-history events (e.g., pregnancies) or diseases (rev. in Tang et al., 2016). Moreover, our high-resolution synchrotron images of the skull reveal the presence of enamel defects in ‘Little Foot’s' teeth. Enamel hypoplasias are indicators of physiological stress experienced during childhood and may be related to diseases or dietary deficiency/nutritional stress (rev. in Guatelli-Steinberg and Huffman, 2012). Interestingly, enamel defects in StW 573’s lower left canine are found at the same distance of the cemento-enamel junction as in other Australopithecus lower canines from Sterkfontein (Guatelli-Steinberg, 2003). Our study thus further confirms that high-resolution synchrotron radiation may reveal very fine details of enamel defects, with substantial differences in the appreciation of hypoplasia between 21.23 and 7.91 microns of spatial resolution.
Additionally, the opening of the diploic vessels in the cranial vault of ‘Little Foot’ could be imaged. Since this structure may be involved in brain thermoregulation, future synchrotron-based 3D virtual reconstruction of the diploic network in early hominin skulls could be particularly useful for determining when and how the complex human-like thermal regulation system emerged (rev. in Bruner, 2017). In particular, such data would contribute to explore the potential correlations between large proportions of diploic bone in Australopithecus and related expansion of the diploic vessels (Beaudet et al., 2018).
To the best of our knowledge, this is the first time that histological features of the compact bone of a Pliocene hominin skull have been non-destructively observed. Since the microscopic organization of the compact bone may have age-related biological significance, the presence, identification, and characterization of such histological structures are of particular interest for determining the age-at-death of fossil specimens (e.g., Ericksen, 1991), particularly in combination with the dental markers discussed above. For example, the branching patterns identified in this study are also found in extant humans (Maggiano et al., 2016). In the sample analyzed here, we could only identify one transverse connection, which confirms the advanced age of StW 573 since this type of branching is more common in young individuals (Maggiano et al., 2016; Clarke and Kuman, 2019). Similarly, the spatial organization of the vascular canal network has been suggested to reflect functional adaptation throughout the individual’s life and/or growth rate in fossil long bones (Ricqlès et al., 2000). Consequently, this histomorphological parameter represents a relevant proxy for evaluating the properties of the loading environment or developmental pattern in fossil specimens (Ricqlès et al., 2000). In our preliminary results, the vascular canals are more abundant close to the trabecular bone in the mandible of StW 573, which may indicate an area of intense bone remodeling, potentially in response of biomechanical loading. The fact that the vascular canals could be successfully reconstructed in 3D in the mandible of a 3.67-million-year-old fossil specimen such as ‘Little Foot’ reveals the invaluable contribution of synchrotron radiation in refining our knowledge of fossil hominin paleobiology at a histological level. For instance, a more comprehensive analysis of the compact bone microstructures should provide new insights into the evolution of the bone modeling/remodeling process, which is a fundamental aspect of bone functional adaptations in the human lineage. We might expect that, because of changes in the biomechanical environments (in the masticatory system but also in relation with locomotory adaptations), the organization of the Haversian system in the skeleton may have varied throughout the hominin lineage.
|
|
|
|
Post by Admin on Mar 12, 2021 23:41:27 GMT
Front. Ecol. Evol., 26 February 2021 | doi.org/10.3389/fevo.2021.639048Visual Depictions of Our Evolutionary Past: A Broad Case Study Concerning the Need for Quantitative Methods of Soft Tissue Reconstruction and Art-Science Collaborations Flip through scientific textbooks illustrating ideas about human evolution or visit any number of museums of natural history and you will notice an abundance of reconstructions attempting to depict the appearance of ancient hominins. Spend some time comparing reconstructions of the same specimen and notice an obvious fact: hominin reconstructions vary in appearance considerably. In this review, we summarize existing methods of reconstruction to analyze this variability. It is argued that variability between hominin reconstructions is likely the result of unreliable reconstruction methods and misinterpretation of available evidence. We also discuss the risk of disseminating erroneous ideas about human evolution through the use of unscientific reconstructions in museums and publications. The role an artist plays is also analyzed and criticized given how the aforementioned reconstructions have become readily accepted to line the halls of even the most trusted institutions. In conclusion, improved reconstruction methods hold promise for the prediction of hominin soft tissues, as well as for disseminating current scientific understandings of human evolution in the future. Introduction: Why Study and Reconstruct Muscles? At a time in which we are increasingly exposed to acclaims about new powerful genetic tool in the media and academia, one may wonder as to why we would focus on muscle reconstructions at all in this introductory paper of this special issue. This is particularly the case since genetic tools are now being used in studies that have been typically done with anatomical tools in the past, such as those concerning phylogenetic reconstructions. Actually, molecular tools are now being used to undertake facial reconstructions, an area that was exclusive to anatomy until very recently. In September 2019, newspapers across the globe reported with astonishment that a new method based on DNA information recovered from the remains of extinct individuals known as the Denisovans enabled scientists to give them a face. Namely, those scientists gleaned anatomical clues from ancient genomes to put together a rough composite portrait of a young female that lived at Denisova Cave in Siberia 75,000 years ago (Gokhman et al., 2019), despite the fact that only small fragments of bones and teeth of Denisovans were found and their skeletal anatomy has not been documented. We will obviously not discuss here the details of that paper and its artistic repercussions, nor the way in which it affected the way Denisovans are perceived by the broader public, although we will briefly refer below to some other similar studies. Rather, the point is that, if we have all these new tools, including eventual facial reconstructions in the future, are anatomical fossil reconstructions destined to become unimportant? The answer is that this is not at all the case; as will be seen in the present paper, and in this special issue as a whole, it is in fact the opposite. There has been a renewed interest in such reconstructions, using new methods and expanding them to tissues other than skeletal ones, such soft tissues like muscles, arteries, veins, and nerves, making them more complete and comprehensive than ever before. This special issue is, in itself, the proof of that, as it would have been difficult to do a whole issue with so many papers from top scholars completely dedicated to muscle reconstructions a few decades ago. In fact, this new interest in fossil muscle reconstructions is part of a resurgence of the study of comparative anatomy per se—the now re-awoken “sleeping beauty”, to paraphrase Virginia Abdala—which was in great part a by-product of the rise of Evo-Devo in the past decades (Diogo, 2018). Some years ago, one of us, with Bernard Wood (Diogo and Wood, 2013), published a paper summarizing why the study of muscles continues to be extremely important for not only Evo-Devo, but also for evolutionary biology, anatomical sciences, biological/physical anthropology, and many other fields. As noted in that paper, a major reason why molecular tools have not yet completely eclipsed anatomical ones in studies of evolutionary relationships is that it is still not possible to recover DNA for most of the millions of species that became extinct much before the time that Denovisans did. For instance, no DNA has been recovered for the fossil taxa that are the central focus of this special issue; those representing the transitions from fishes and early tetrapods. Therefore, phylogenetic works of such groups have been traditionally done mainly with bones but are also increasingly using soft tissues—particularly muscles as will be seen in this issue. One of the reasons for this is, as noted in that paper, studies by us and various other authors on the whole osteichthyan clade (bony “fish” plus tetrapods), and on specific groups such as our own (primates), have shown that although osteological structures often provide more potential characters for phylogenetic analyses, myological characters tend to be more useful for inferring the phylogenetic relationships among higher clades. Indeed, this seems to apply even to fossil taxa such as non-avian dinosaurs (e.g., Dilkes, 2000). This therefore illustrates how crucial it is to undertake accurate muscle reconstructions of fossils, to not only understand their functional morphology, and biology as a whole—bones do not move without muscles—but also to learn more about their evolutionary relationships, history, and adaptations. This is moreover crucial, as will be discussed below, for science dissemination and the way the broader public perceives those fossil taxa, such as early tetrapods, dinosaurs, and even the closest extinct relatives of the human lineage. We are thus living in a fascinating time in which instead of a decrease of interest in muscles, there is an exponential interest in developing new tools and ways to reconstruct them more accurately in fossil taxa, and in displaying them artistically in the web, dissemination books, popular movies and documentaries, and museum fossil displays. Due to the particular interest in the reconstructions of fossils of our human lineage for all these types of media, their artistic repercussions, and the way they influence the public perception and narratives built around them—including, unfortunately, racist and misogynistic ones, as shown in Moser’s (1996) book Ancestral images: the iconography of human origins—in this introductory paper we will focus on our own lineage. The idea is to show that the focus of this issue, muscle reconstructions, has not only scientific repercussions, but also societal and artistic implications. As will be shown in sections below, such reconstructions involve major complexities and difficulties, but also bring fascinating new opportunities. Over the last century, there has been a huge interest in reconstructing the face of members of our human lineage that lived many thousands, or even some millions, years ago. However, most of these are based on unfalsifiable ad hoc stories that have little or no empirical evidence. For instance, it has been said that the prognathic faces of Australopithecus were more similar to our closest living relatives, the great apes (chimpanzees, gorillas, and orangutans), than to anatomically modern humans. Based on this observed similarity, some have assumed that the soft tissues covering their faces would also have been more similar to those of apes than to those of Homo sapiens (Aiello and Dean, 1990; Gurche, 2013). This kind of rhetoric, which is largely untestable, is frequently deployed in the process of reconstructing Plio-Pleistocene hominins (N.B., in this paper hominins means all humans since we split from common ancestors with separately evolving lineages). It is based on a kind of interpretation called retrodiction, which is an intuitive method for predicting the past based on present observations of natural phenomena. It is based on Charles Lyell’s uniformitarian principle underlying the entire evolutionary science. But how reliable is retrodiction? Could not this rhetoric be questioned? Here, we review the practice of hominin reconstruction from a scientific perspective and address some of its broader implications. Specifically, we begin by presenting some of the earliest examples of hominin reconstruction followed by a review of the current methods used. We then show where future research holds promise for improving existing methods and producing scientifically accurate reconstructions, followed by a discussion of our own view on the ethical and societal implications of artistic interpretations of hominins. Our aim is to identify areas where fresh research is needed, which can be applied to other non-human or non-primate taxa. Our fascination with hominin reconstructions—and the basis for this review—stems chiefly from the work carried out by two of us (RC and GV) over the last 6 years attempting to reproduce 3D reconstructions of extinct hominins often using the muscle data that have been recently made available for apes by another co-author (RD) and his colleagues. Although many 2D reconstructions of hominins exists, which are arguably just as important as 3D reconstructions, we will focus mainly on 3D reconstructions as these are the ones that we have spent the most time trying to replicate. It is hoped that including our own reconstructions in this review will help to expose the limitations of existing methods and to substantiate our claim that the practice is lacking a robust scientific and empirical foundation. As we shall show, many of the questions regarding the appearance of Plio-Pleistocene hominins are yet to be answered and most, if not all, reconstructions are based on methods that are irreplicable. This once again highlights the difficulties and complexities of muscle reconstructions but also the enormous opportunities that we now have to make progress in the area of muscle, facial, and whole-body reconstructions. |
|
|
|
Post by Admin on Mar 13, 2021 0:27:50 GMT
A Brief History of Hominin Facial Reconstruction
The earliest reconstructions of hominins were carried out in the late nineteenth and early twentieth centuries by artists and scientists in the form of both 2D and 3D portraits as well as whole-body reconstructions, produced soon after the discovery of various fossils. As very few hominin fossils have ever been found—it is, after all, a well-known fact that there are more active physical anthropologists today than there are hominin finds—it is relatively easy to compare reconstructions of the same individual. As we shall show, there are only a handful of well-preserved skulls suitable for reconstruction, which not only makes it easy to compare appearances between reconstructions of the same individuals produced by separate practitioners, but also highlights the role of how individually constructed knowledge about human evolution can affect their results. We would like to be transparent with the reader and admit that this section is by no means a complete list of all of the reconstructions that have ever been produced, however, it does include the most well-recognized practitioners and reconstructions that were featured in scholarly publications, scientific textbooks, and on display at institutions of international repute.
The best documented 3D hominin reconstructions based on scientific methods were produced by the Russian anthropologist and archeologist Mikhail Gerasimov (Gerasimov, 1971). Gerasimov is especially renowned for his contributions to the field of forensic facial reconstruction—now more commonly referred to as facial approximation—which is the process of reproducing a likeness that can assist in identifying an individual from a skull found in a forensic context. In his published work, Gerasimov used his forensic methods—for a review of these methods, see Ullrich and Stephan (2016)—to reconstruct two Australopithecines as well as various members of the genus Homo. The best known 3D reconstructions of hominins today are produced by John Gurche (Balter, 2009; Gurche, 2013). Gurche has allegedly reconstructed over fifteen hominin individuals that are featured in the Smithsonian National Museum of Natural History in Washington, D.C. These reconstructions include Sahelanthropus tchadensis, Australopithecus afarensis, Australopithecus africanus, and Paranthropus boisei. Gurche has also reconstructed individuals from the genus Homo, including Homo erectus, Homo heidelbergensis, a Neandertal, and LB1 (Balter, 2009; Gurche, 2013). Other well-known practitioners of 3D reconstruction include Élizabeth Daynès, Gary Sawyer, Viktor Deak, Philippe Froesch, and Adrie and Alfons Kennis (Balter, 2009).
Is it important to note here that not all reconstructions of hominins have been produced in 3D, 2D reconstructions are arguably more numerous and thus any review would be incomplete without acknowledging them. In general, 2D reconstructions appear to conform less to the scientific approach and more to artistic intuition but this fact does not weaken their power of influence on public perceptions about human evolution and are therefore relevant to this review. Zdeněk Burian is one of the most celebrated 2D paleoartists in physical anthropology and produced a number of illustrations of hominins depicted in their ancestral environments (Jelínek, 1975). Jay Matternes also produced 2D reconstructions. One of these illustrations is of an individual of Australopithecus afarensis and is regarded by world-renowned paleoanthropologist Donald Johanson—who was consulted during the production of this reconstruction—as one of the “finest representations of this species” (Johanson, 1981). With respect to Burian, little is known regarding how the soft tissues were extrapolated from the fragmentary fossils upon which his reconstructions were based. Here we can only assume that these illustrations were reconstructed intuitively. In contrast to Burian, Matternes provides a full description of his methods. The reconstruction, he says, was made over an image of a composite reconstruction of an Australopithecus afarensis skull (Kimbel et al., 1984; Kimbel and White, 1988). The masticatory muscles and muscles of expression were constructed over the skull first, then existing methods for approximating the other features of the face were borrowed from the facial approximation literature, including mouth width determination, locating the eyeballs within the eye sockets, as well as deciding on the ear morphology, flexure wrinkles, and hirsuteness (Johanson, 1981).
Anyone attempting to reconstruct a hominin ought to be aware of the aforementioned practitioners and their influence on the current state of the practice. Scientists like Gerasimov and artists like Burian were some of the first to attempt to produce a hominin face from skeletal remains. Their results have functioned as hypotheses for the facial appearances of their subjects and while not all of these hypotheses may appear equally valid to the reader, we would like to propose that in the absence of a well-established systematic approach for reconstructing hominin soft tissues, these works provide valuable insights into each practitioners’ methodology. However, although these works have helped immensely in encouraging interest in human evolution, the methods employed by the aforementioned practitioners remain largely unchanged today. Gerasimov’s methods have seen no improvement in their application to hominins and Burian’s artistic intuition has been replicated by other artists, such as the paleoartist Mauricio Antón, with varying results.
The Problem of Variability
Differences among hominin reconstructions were first systematically documented in a pivotal study by Karen Anderson, in which 860 hominin reconstructions were assessed from 55 museum displays across Europe and Australia. Inconsistencies between reconstructions of the same individual were found in both their surface appearances and body proportions (Anderson, 2011). To make matters worse, most hominin reconstructions were found to be presented without any rigorous empirical justifications. Despite this, and to the surprise of the authors, the same reconstructions are commonly cited in the scientific literature and presented in scientific textbooks on human evolution (Jelínek, 1975; Balter, 2009; Jablonski, 2013; Roberts, 2018). So severe are the differences between reconstructions of the same individual that it is almost as though the practitioners had never encountered another hominin reconstruction before commencing their own. From a scientific point of view, there are only two ways of explaining an error of this magnitude: either (1) the reconstructions are purely artistic interpretations based on individually constructed knowledge about human evolution, which can vary between practitioners and ultimately results in variability, and/or (2) the practitioners were using unreliable reconstruction methods. Why such varying reconstructions continue to be used in the dissemination of science when such reconstructions have never been formally verified is disconcerting to us because the quality of knowledge perpetuated by their use is clearly inconsistent. To make matters worse, consider the reconstruction of Lucy presented at the “Answers in Genesis” ministry’s Creation Museum in Petersburg, Kentucky. While Lucy was indeed a primate, the decision to reconstruct this specimen as a knuckle-walker is an obvious error. However, the argument of variability put forward by the Creation Museum is a valid one that has, as of yet, not been addressed by the scientific community.
To the knowledge of the authors, Gerasimov is the only practitioner to express doubt about the use of his methods for reconstructing the faces of ancient hominins. He acknowledged from the outset that there was an inherent risk in interpolating soft tissue depth data collected from orangutans into his reconstruction of the Australopithecus africanus specimen Sts 5 (Gerasimov, 1971). In contrast, Gurche is on record saying that he developed his method for reconstructing hominins from personal research carrying out dissections of extant apes and modern humans (Gurche, 2013), but this research has never been formally verified nor published in any scientific literature. Regarding Élizabeth Daynès, Gary Sawyer, Viktor Deak, and Adrie and Alfons Kennis, none of these practitioners have ever published any details regarding their methods or justifying their results. Thus, at present it is evident that hominin reconstruction is a practice lacking a robust scientific and empirical foundation.
Methods and Techniques Used for Reconstructing Hominins
To explore the question of why the aforementioned variability has and is still occurring, we will evaluate the evidence and methods available to practitioners of hominin reconstruction. As stated in the Introduction, to aid in our review we will present the various reconstructions performed by RC and GV over the last 6 years as case studies to (1) exemplify the quality of evidence that is available in each case and (2) to show what existing methods were employed in each case to explore their strengths and weaknesses.
|
|

