|
|
Post by Admin on Jul 10, 2019 17:59:16 GMT
Morphometric analysis The results of the principal components analysis (PCA) were similar to those described previously for similar neurocranial datasets analyzed by one of us (KH; Figure 3; [6]–[8]). The first principal component accounted for 32.4% of the total variance and separated archaic from modern human specimens. The two H. erectus (s.l.) specimens fell at the extreme negative of this axis, followed by Neanderthals and H. heidelbergensis (s.l.). Modern human populations were characterized by more positive scores on PC 1, and there was only minimal overlap among their 95% confidence ellipses and that of the Neanderthals. The Middle-Late Pleistocene African specimens (LH 18, Singa, Djebel Irhoud 1 and 2) and the early modern human specimens from Qafzeh and Skhul fell in the intermediate zone between Neanderthals / H. heidelbergensis on one hand and modern humans on the other. Qafzeh 9 was the exception, falling on the positive end of PC 1 and close to Upper Paleolithic European specimens. The latter sample, which included some of the earliest modern human specimens in Europe (Mladeč 1 and 5, Oase 2, Muierii 1, Cioclovina), clustered within the modern human range of variation, and not in the zone of overlap with the archaic specimens. One of the two specimens from Zhoukoudian Upper Cave (UC 101) had a more negative PC 1 score similar to that of Qafzeh 6 and Jebel Irhoud 2 (see also [8]). Iwo Eleru showed a similarly negative PC 1 score, falling closest to LH 18, Saldanha (Elandsfontein) and Spy 2 along this axis. PC 1 reflected differences in the shape of the neurocranium from an elongated, low vault and large, evenly thick, supraorbital torus to a rounded, antero-posteriorly shorter vault, and thinner supraorbital torus with differentiated medial and lateral segments.  Figure 3. Results of the multivariate statistical analysis of landmarks and semilandmarks. Top: Principal components analysis, PC1 and 2. Cranial shape differences along PC 1 are shown below the graph. The top graph shows a Minimum Spanning Tree of the Inter-individual Procrustes distances for the fossil specimens (black lines connecting specimens). Specimen labels as in Table 1. Bottom: Canonical variates analysis, CV 1 and 2. Symbols: Grey diamonds. Modern humans; Black up triangles: Upper Paleolithic modern humans; Purple up triangles: Late Pleistocene African and Near Eastern hominins; Red stars: H. neanderthalensis; Red squares: H. heidelbergensis (s.l.); Black squares: H. erectus (s.l.). Ellipses indicate 95% confidence ellipses for Neanderthals (red) and modern humans (gray). PC 2 (11.3% of the total variance, Figure 3) appeared to reflect variation among modern humans, with the sub-Saharan African, the Khoisan, Oceanic and Upper Paleolithic samples clustering around zero, and on the positive end of this axis. The Andaman sample was restricted to the negative side, while the Inuit fell around zero. The Europeans, N. Easterners, Asians and Iberomaurusians spanned the entire length of PC 2. The LPA sample was also quite spread out along this axis, while the Neanderthals, H. erectus and H. heidelbergensis s.l. were relatively centered around zero and on the negative side. Iwo Eleru was once more placed closest to LH 18 and Saldanha. Crania with relatively negative scores on PC 2 displayed antero-posteriorly short and medio-laterally wide shapes, while those with positive scores showed antero-posteriorly long and medio-laterally narrow vaults.  Figure 4. Shape comparisons of Iwo Eleru. Iwo Eleru (black) compared to the modern human mean configuration (A, gray), to the Late-Middle Pleistocene African mean configuration (B, gray), and to its closest modern human neighbor in overall shape (C, gray). Iwo Eleru generally showed large distances to the other specimens in this analysis. It displayed the shortest inter-individual Procrustes distance to LH18 (0.080), and next closest to La Chapelle-aux-Saints (0.087) and a recent Australian (0.089). LH18 itself was closest to the Middle Pleistocene Saldanha individual (0.059). It also showed relatively small Procrustes distances to two Upper Paleolithic Europeans, Oase 2 (0.071) and Predmostí 3 (0.074). La Chapelle showed the shortest distances to other Neanderthals: Guattari (0.054), La Quina 5 (0.054), Spy 1 (0.057), Amud 1 (0.061), La Ferrassie 1 (0.067), Feldhofer 1 (0.075), Shanidar 1 (0.076); as well as to older specimens: Sima de los Huesos Cranium 5 (0.069), Dali (0.060), Irhoud 1 (0.067). The Procrustes inter-individual distances were used to generate a minimum spanning tree for the fossil specimens on the plot of PC 1 and 2 (Figure 3). The results of the canonical variates analysis (CVA) were consistent with those of the PCA (Figure 3). The first canonical axis (49.7%) separated archaic from modern specimens, with late archaic or early modern humans (the Irhoud specimens, Qafzeh 6, Singa, LH18) generally falling in an intermediate position. Iwo Eleru, as well as Upper Cave 101, also fell in this region, with the former and LH18 being just at the outskirts of the Neanderthal confidence ellipse. The Mahalanobis squared distances among the predefined groups are reported in Table S2. Iwo Eleru showed large distances from all other groups. The smallest distance was to the Upper Cave specimens, themselves a very small group of just two individuals. Relatively small squared distances were also shown between Iwo Eleru and the Qafzeh-Skull group, Neanderthals and H. erectus (s.l.). The departure of Iwo Eleru from the modern human average cranial shape was further underlined by the comparison of its landmark configuration to the mean configuration of modern humans (Figure 4A). Iwo Eleru was characterized by a more elongated cranial vault and flattened frontal and parietal bones. Its browridge was also slightly more forward projecting than the average modern human shape. Iwo Eleru was more comparable to the mean LPA landmark configuration in its elongated and low cranial shape and the degree of browridge projection (Figure 4B). Its nearest recent human neighbor, an Australian female (Figure 4C), also showed a relatively low vault and pronounced browridges. However, the latter specimen exhibited an overall more curved sagittal profile than Iwo Eleru, with a more steeply rising frontal bone, an expanded and more curved parietal and a more rounded occipital with a lower position of inion, all typical modern human conditions. |
|
|
|
Post by Admin on Jul 10, 2019 20:31:17 GMT
Uranium-Series Dating The elemental U and Th values vary greatly along the profiles of the bone sample from the Iwo Eleru skeleton (left-hand diagrams in Figures S1 and S2). The outer surfaces and pores show measureable Th concentrations, which indicate the presence of detrital material. There is a gradient of higher U-concentrations in the centre of the bone (around 80 to 100 ppm) towards the outsides where U-concentrations of 50 to 60 ppm were measured. These lower U-concentrations may either be due to a denser bone structure, with less internal surface area for adsorbing uranium, or to leaching of U from the volumes close to the surface. The apparent U-series age estimates vary between about 10 to 14 ka in the central part of the bone and increase in some laser scans to between about 15 to 17 ka closer to the surface. The right-hand columns of Figures S1 and S2 show the plot of apparent U-series age versus U-concentration. It is expected from the diffusion-adsorption model for U-uptake [9]–[12] that spatially resolved analyses across a homogeneous bone yield u-shaped or constant U-concentration and apparent U-series age profiles. In ideal circumstances, a plot of apparent U-series age versus U-concentration would either be flat or show increasing U-series age estimates with increasing U-concentrations. This is clearly not the case for the results on Iwo Eleru (Figures S1 and S2, right hand columns). The apparent U-series age estimates are more or less constant in the inner part of the bone fragment and increase towards the outside while the U-concentrations decrease. One of the problems of U-series dating of bone is that the different domains within a bone may give very different U-series ages. The best age estimate is actually not derived by simply averaging all results, but from identifying the domains that had experienced the fastest U-uptake and have remained a closed system since. This may only apply to very small volumes (see [13]). The U-series analyses of the central part of the bone give a minimum age of the sample. Excluding any results with more than 10 ppb Th (indication of contamination) and U concentrations of less than 75 ppm, all scans give compatible results with mean ages ranging between 10.6±1.7 (Track 2) and 12.7±1.6 ka (Track 4), see right hand columns in Figures S1 and S2. Most apparent U-series age results from the central parts of the bone are within 9 and 14 ka. Virtually all age results overlap within their errors. Most of the scatter is likely due to statistical variation. The best age estimate is thus derived from the mean value of all Th-free age estimates in the central part of the bone (11.7±1.7 ka). This age estimate is derived from more than 80% of the exposed bone area (excluding the pores), which may be taken as an indication that most of the uranium was accumulated within a relatively short time range. The average U-series age increases by less than 1 ka (i.e. less than 10%) by including the Th-free results of the domains with lower U-concentrations. Considering that the outer U-concentrations are lower by up to a factor of four compared to the interior part, U-leaching is apparently not the main cause for the reduced U-concentrations. The question is whether the specimen could be substantially older. Most older apparent age estimates are associated with relatively high Th and low U concentrations, either close to the outside or in pores (Tracks 1, 3, 4, 11, and 12). Here, the older apparent U-series results may well have been influenced by detrital material as well as U-leaching. Other older results (>15 ka) are seen for example in Track 8 (around cycle 3100), Track 11 (around cycle 430 and 550) and Track 12 (around cycle 1480), see arrows in the respective diagrams in Figure S2. In Track 12, the section between cycle 1481 and 1488 yields an age of 19.3±1.8 ka. The immediately adjacent sections yield 13.9±2.6 ka (cycles 1472 to 1481) and 10.8±2.1 ka (cycles 1488 to 1505). It is a simple statistical fact that some subsamples of a single population will deviate from the other results by more than 2-σ. The most likely apparent age result comes from the average of this section. The same seems to apply to the other marked sections. Nevertheless, Tracks 9 (between cycles 3076 and 3161) and 11 (between cycles 726 and 791, see circles in the respective diagrams of Figure S2) show wider domains with relatively old apparent U-series ages (15.0±1.3 and 16.3±0.5 ka), which do not depend on the U-variations within these domains. While U-leaching cannot entirely be excluded in these sections, the lower U-concentrations may just as well be due to different amounts of internal surface area. These two sections are close to the lower surface and could have preserved the original U-series isotope signatures. If this is the case, the most likely age of the bone is around 16.3 ka. In summary, the minimum age of the bone is around 11.7±1.7 ka while some domains indicate an age as old as 16.3±0.5 ka.  During a recent examination of our morphological 3-D coordinate dataset, it was ascertained that it included a subadult specimen not listed in the publication. This specimen was not shown in the plots but mistakenly remained in the analysis as part of the early modern human sample (EAM). We thank Dr. Waddell for bringing this issue to our attention. We have re-analyzed our data excluding this specimen. Although some of the numerical results change slightly, the main findings, relationships among samples and our conclusions do not change. The PCA and CVA plots, as well as the minimum spanning tree, are nearly identical to those originally reported (Corrected Figure 3). The new Mahalanobis D2 results are similar in the values obtained and pattern of relationships among samples to the ones originally reported (Corrected SI Table 2). The main difference is the distance between Iwo Eleru and the early modern human Qafzeh-Skhul sample from the Levant (EAM), which dates to between 90-130 ka, in the distances corrected for unequal sample sizes. This group, and not the much later Zhoukoudian Upper Cave sample as originally reported, is the closest neighbor to Iwo Eleru in the corrected Mahalanobis D2. This result, although different from that originally reported, strengthens our conclusion that Iwo Eleru bears greater morphological similarity to early rather than late modern human samples from Africa and Eurasia. The revised dataset and the updated key for the labels used in the dataset are also provided with this Correction. Figure 3: Discussion Our analysis indicates that Iwo Eleru possesses neurocranial morphology intermediate in shape between archaic hominins (Neanderthals and Homo erectus) and modern humans. This morphology is outside the range of modern human variability in the PCA and CVA analyses, and is most similar to that shown by LPA individuals from Africa and the early anatomically modern specimens from Skhul and Qafzeh. Iwo Eleru is distinct from the recent African samples used here (although the range of recent modern human variation encompasses relatively low and elongated cranial shapes approaching this condition). Past work has suggested that neurocranial shape reflects population history relatively reliably among modern human populations [14], [15]. Although we did not find unambiguous strong affinities between Iwo Eleru and the samples used here, its overall morphological similarities with early modern humans suggest a link to these early populations and possibly a late Middle-early Late Pleistocene chronology. Nonetheless, the archaeological setting, stratigraphy, previous radiocarbon [see 4] and our new U-series dating indicate a much younger, terminal Pleistocene age for this cranium. Such a late chronology for the Iwo Eleru cranium implies that the transition to anatomical modernity in Africa was more complicated than previously thought, with late survival of “archaic” features and possibly deep population substructure in Africa during this time. Thus our restudy of the Iwo Eleru cranium confirms previously noted archaic cranial shape aspects, and the U-series age estimates on its skeleton support the previously proposed terminal Pleistocene date for this burial. Our findings also support suggestions of deep population substructure in Africa and a complex evolutionary process for the origin of modern humans [16], [17], [7], [18], [19], [20], [21]. Perhaps most importantly, our analysis highlights the dearth of hominin finds from West Africa, and underscores our real lack of knowledge of human evolution in that region, as well as others. As also indicated by restudy of the Ishango (Congo) fossils [22], Later Stone Age fossils from at least two regions of Africa retain significant archaic aspects in their skeletons. We hope that the next stage of this research will extend studies to the Iwo Eleru mandible and postcrania, and to comparative materials such as those from Ishango. Harvati K, Stringer C, Grün R, Aubert M, Allsworth-Jones P, Folorunso CA (2011) The Later Stone Age Calvaria from Iwo Eleru, Nigeria: Morphology and Chronology. PLoS ONE 6(9): e24024. |
|
|
|
Post by Admin on Jul 12, 2019 8:51:14 GMT
The Apidima Cave complex was excavated in the late 1970s. Two partial crania (skulls without the lower jaw), named Apidima 1 and Apidima 2, were recovered in a single block of a type of rock called breccia. Neither fossil was previously described in detail. Apidima 2 includes the facial region of the skull and had been identified as a Neanderthal7. Apidima 1 consists of only the back of the skull and had not been previously allocated definitively to a species. Harvati and colleagues used computed tomography to scan the fossils, and generated a 3D virtual reconstruction of each specimen. They analysed each fossil to assess aspects of its shape, and thus to determine the fossils’ similarity to those of other species.  Apidima 2 is badly damaged owing to previous breakage and distortion. Analyses of all four generated reconstructions of the fossil were consistent with it being an early Neanderthal. Apidima 1 is also damaged, but the specimen is not too badly distorted, so mirroring its right and left sides yielded a good reconstruction. The authors’ extensive comparative analysis indicates that this fossil is an early member of H. sapiens. The posterior part of the cranium is rounded like that of H. sapiens, and it lacks classic Neanderthal features, such as the distinctive occipital ‘chignon’ — a bulge at the back of the skull that is shaped like hair tied in a bun. Earlier dating8 of a fragment of Apidima 2 using a method called uranium-series analysis indicated a minimum age of around 160,000 years. Harvati and colleagues report a more extensive set of uranium-series dating analyses, which surprisingly reveal that Apidima 1 and Apidima 2 are of different ages, even though they were found in close proximity. Apidima 2 is around 170,000 years old — well within the age range of other Neanderthal fossils found across Europe (Fig. 1). Apidima 1 is dated to be at least 210,000 years old, which is much older than any other widely accepted H. sapiens fossils found outside Africa. 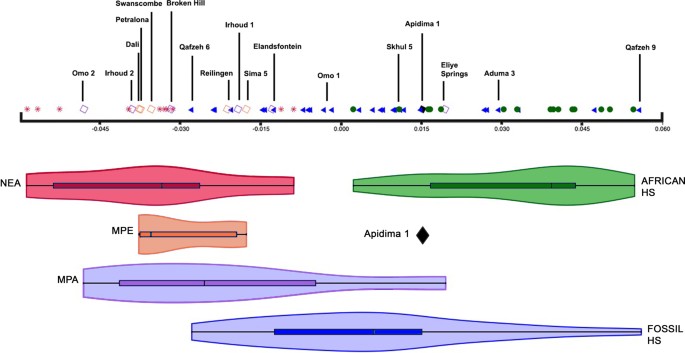 This finding reveals that at least two species of hominin (humans and human relatives from the branch of the family tree after our split from chimpanzees) inhabited southeastern Europe approximately 200,000 years ago. The discovery of an H. sapiens fossil in Apidima raises questions about what happened to this population. Given that this H. sapiens existed at a time when there is substantial evidence for a Neanderthal presence at other European sites, was it part of a population that was unable to compete successfully with Neanderthals, especially in the unstable climate of that time? Perhaps one or more times, the two species replaced each other as the main hominin group present in this region. Such patterns of replacement characterize the distribution of modern humans and Neanderthals in the Levant region of the Middle East between 250,000 and 40,000 years ago. Homo sapiens replaced Neanderthals across Europe between approximately 45,000 and 35,000 years ago6, eventually giving rise to the ancestral population of Europeans alive today1. This evidence from Apidima, along with other discoveries, demonstrates that, on more than one occasion, modern humans kept pushing north and westwards from Africa and the Levant into Europe. Rather than a single exit of hominins from Africa to populate Eurasia, there must have been several dispersals, some of which did not result in permanent occupations by these hominins and their descendants. 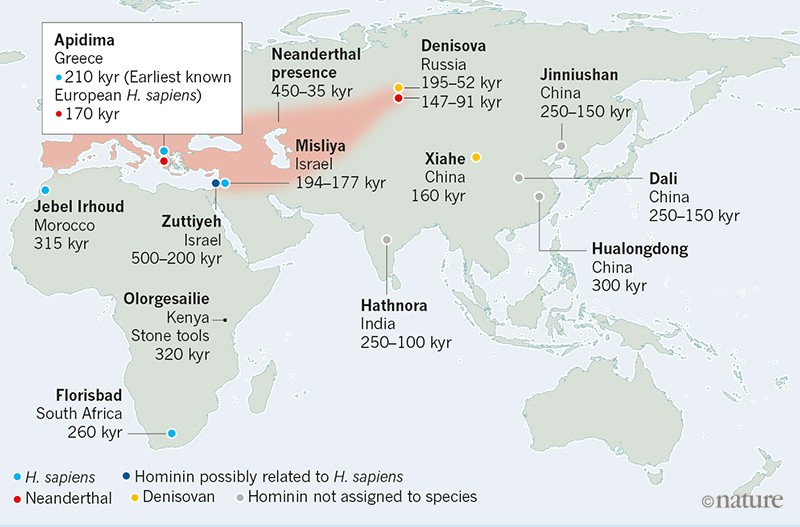 Figure 1 | Some key early fossils of Homo sapiens and related species in Africa and Eurasia. Harvati et al.5 present their analyses of two fossil skulls from Apidima Cave in Greece. They report that the fossil Apidima 1 is an H. sapiens specimen that is at least 210,000 years old, from a time when Neanderthals occupied many European sites. It is the earliest known example of H. sapiens in Europe, and is at least 160,000 years older than the next oldest H. sapiens fossils found in Europe6 (not shown). Harvati and colleagues confirm that, as previously reported7, Apidima 2 is a Neanderthal specimen, and they estimate that it is at least 170,000 years old. The authors’ findings, along with other discoveries of which a selection is shown here, shed light on the timing and locations of early successful and failed dispersals out of Africa of hominins (modern humans and other human relatives, such as Neanderthals and Denisovans). kyr, thousand years old. A jaw of an early modern human from Misliya Cave in Israel has been dated to approximately 194,000–177,000 years ago11. Other early modern human fossils have been found at Skhul and Qafzeh in Israel, dated to around 130,000–90,000 years ago12. All of these early Eurasian human fossils seem to represent what might be called ‘failed’ dispersals from Africa — they reached the Middle East and southeastern Europe, but did not persist in these regions. There is evidence that these populations were replaced at these or neighbouring sites by Neanderthals. |
|
|
|
Post by Admin on Jul 18, 2019 17:50:47 GMT
 When modern humans started emerging from Africa and spreading throughout Eurasia, they found many places already occupied by older hominids such as Neanderthals and Denisovans. As humans do, we got rather friendly with our new neighbours: evidence of that hanky panky lives on in our DNA today. After closely analysing the existing literature, Teixeira and his colleague biologist Alan Cooper have identified two such 'ghost' ancestors in modern DNA. The first, identified in Eurasian DNA with the help of artificial intelligence, was widely reported earlier this year. The second, however, was reported last year, a detail that flew under the radar in a larger paper: a mysterious, and inconclusive, genetic signature exclusively found in the population of Flores, Indonesia. It appears to be as divergent from modern human DNA as Neanderthal or Denisovan DNA is. 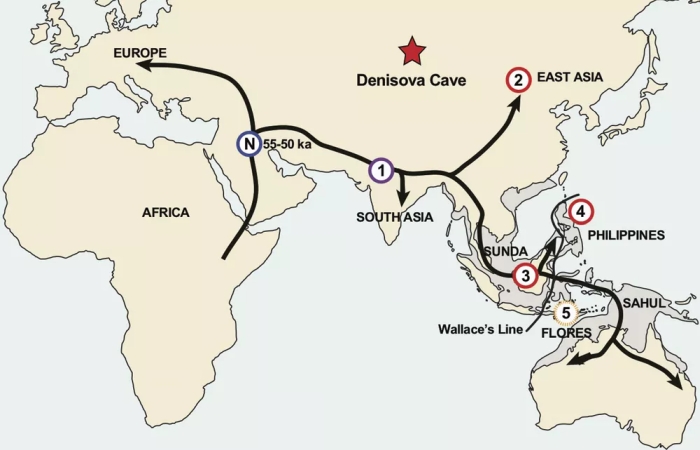 As the modern humans moved farther east, across into islands of Southeast Asia, they seem to have run into more groups. "At least three other archaic human groups appear to have occupied the area, and the ancestors of modern humans mixed with them before the archaic humans became extinct," Teixeira said.nOne of those groups was the Denisovans. The other two remain a mystery. The first unknown extinct hominid - named EH1 - was roughly genetically equidistant from Denisovans and Neanderthals. The ancestor of all Asian and Australo-Papuan populations bred with EH1, resulting in 2.6 to 3.4 percent shared EH1 ancestry.  It's less strong now, but that genetic signal can still be detected in the DNA of Aboriginal Australians, East Asians and Andaman Islanders. This led the researchers to tentatively conclude that EH1 likely occupied a region in northern India, where a group of modern humans - the migration branch that went on to Asia, Australia and the Papuan islands - encountered them (1 on the map above). Modern humans also seemed to have interbred with Denisovans in a number of locations, such as East Asia, the Sunda Shelf, and the Philippines (2, 3, and 4 on the map). Evidence for EH2 - the extinct hominid that interbred with modern humans on Flores - is a little less clear. It only appears in short-statured people that live near Liang Bua Cave - where Homo floriensis was discovered. So it's highly localised, and has somehow remained contained for the roughly 50,000 years since the two groups met (5 on the map).  |
|
|
|
Post by Admin on Aug 29, 2019 7:21:13 GMT
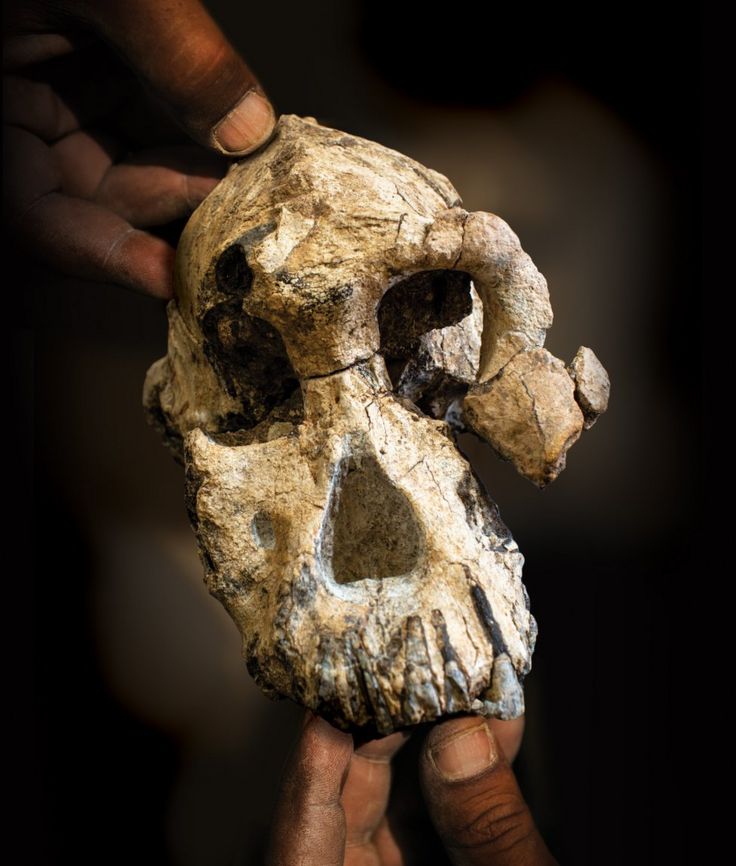 The researchers who discovered the skull say it belongs to a species called Australopithecus anamensis, and it gives researchers their first good look at the face of this hominin. This species was thought to precede Lucy’s species, Australopithecus afarensis. But features of the latest find now suggest that A. anamensis shared the prehistoric Ethiopian landscape with Lucy’s species, for at least 100,000 years, the researchers say. This hints that the early hominin evolutionary tree was more complicated than scientists had thought — but other researchers say the evidence isn’t yet conclusive. 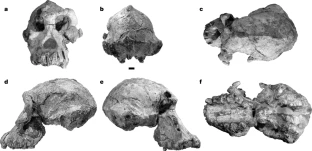 Exceptionally preserved A. afarensis lived in East Africa between about 4 million and 3 million years ago. It is important to the understanding of human evolution because it might have been the ape-like species from which the ‘true’ human genus, Homo, evolved about 2.8 million years ago. Over the past few decades, researchers have discovered dozens of fragments of australopithecine fossils in Ethiopia and Kenya that date back more than 4 million years. Most researchers think these older fossils belong to the earlier species, A. anamensis. It’s generally thought that A. anamensis gradually morphed into A. afarensis, implying that the two species never coexisted. The 3.8-million-year-old hominin skull, discovered at a site called Woranso-Mille in Ethiopia, now suggests otherwise. A team of palaeoanthropologists led by Yohannes Haile-Selassie at the Cleveland Museum of Natural History in Ohio discovered the specimen — called the MRD cranium — in 2016. 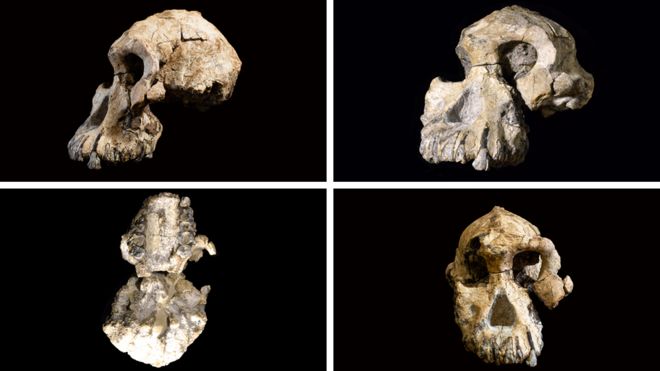 It had been thought that A. anamensis was the direct ancestor of a later, more advanced species called Australopithecus afarensis, which in turn has been considered a direct ancestor of the first early humans in the grouping, or genus, known as Homo, and which includes all humans alive today. The reason for this likely elevated status is because we can now say that anamensis and afarensis actually overlapped in time. The former did not evolve directly into the latter in a neat linear manner, as previously supposed. The realisation comes about through the reinterpretation that the new fossil brings to bear on a previously discovered 3.9-million-year-old skull fragment. That fragment had been assigned to anamensis. Scientists can now see it is actually the remains of an afarensis, pushing this species' origin deeper into the past.  Coexistence? The find makes it possible to reassess other australopith facial fossils, including a 3.9-million-year-old forehead fragment found in Ethiopia in the 1980s. Until now it wasn’t clear whether this fragment belonged to A. afarensis or A. anamensis. But now the researchers have been able to show that the fragment has features that are seen in younger A. afarensis skulls but are absent in the MRD cranium. Assuming the forehead fragment belonged to an A. afarensis individual, and the MRD cranium to an A. anamensis individual, the researchers suggest that the two species may have coexisted in the region for 100,000 years or more. Haile-Selassie and his colleagues say it’s still likely that Lucy’s species evolved from A. anamensis. But they think it did so through a ‘speciation event’: perhaps a small group of A. anamensis became genetically isolated from the general population and evolved into A. afarensis, which eventually outcompeted the wider A. anamensis population. 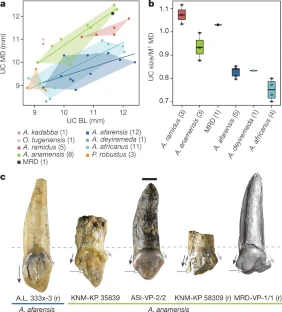 Cranium Specimen, Australopithecus anamensis. A near-complete skull of the species Australopithecus anamensis was discovered in Ethiopia in 2016.Credit: Dale Omori/Cleveland Museum of Natural History An ancient face is shedding new light on our earliest ancestors. Archaeologists have discovered a 3.8-million-year-old hominin skull in Ethiopia — a rare and remarkably complete specimen that could change what we know about the origins of one of humanity’s most famous ancestors, Lucy. 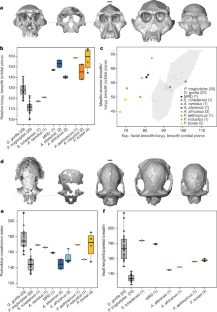 Coexistence? The find makes it possible to reassess other australopith facial fossils, including a 3.9-million-year-old forehead fragment found in Ethiopia in the 1980s. Until now it wasn’t clear whether this fragment belonged to A. afarensis or A. anamensis. But now the researchers have been able to show that the fragment has features that are seen in younger A. afarensis skulls but are absent in the MRD cranium. Assuming the forehead fragment belonged to an A. afarensis individual, and the MRD cranium to an A. anamensis individual, the researchers suggest that the two species may have coexisted in the region for 100,000 years or more. Haile-Selassie and his colleagues say it’s still likely that Lucy’s species evolved from A. anamensis. But they think it did so through a ‘speciation event’: perhaps a small group of A. anamensis became genetically isolated from the general population and evolved into A. afarensis, which eventually outcompeted the wider A. anamensis population. Oldest Homo sapiens fossil claim rewrites our species' history Melillo admits that arguing for a local speciation event rather than the gradual transformation of the entire population might seem like splitting hairs, but she says that understanding exactly how hominin species evolved is a crucial first step to unravelling why they evolved the way they did. Some researchers are ready to consider the possibility that A. afarensis and A. anamensis coexisted. “It’s a very interesting claim,” says David Strait, a palaeoanthropologist at Washington University in St Louis, Missouri. But both Strait and Ward think the evidence isn’t conclusive yet, because it rests heavily on just two fossils — the MRD cranium and the forehead fragment discovered in the 1980s. Strait thinks future fossil finds might help to firm up the idea. Abstract The cranial morphology of the earliest known hominins in the genus Australopithecus remains unclear. The oldest species in this genus (Australopithecus anamensis, specimens of which have been dated to 4.2–3.9 million years ago) is known primarily from jaws and teeth, whereas younger species (dated to 3.5–2.0 million years ago) are typically represented by multiple skulls. Here we describe a nearly complete hominin cranium from Woranso-Mille (Ethiopia) that we date to 3.8 million years ago. We assign this cranium to A. anamensis on the basis of the taxonomically and phylogenetically informative morphology of the canine, maxilla and temporal bone. This specimen thus provides the first glimpse of the entire craniofacial morphology of the earliest known members of the genus Australopithecus. We further demonstrate that A. anamensis and Australopithecus afarensis differ more than previously recognized and that these two species overlapped for at least 100,000 years—contradicting the widely accepted hypothesis of anagenesis. Nature (2019) |
|

