Post by Admin on Mar 5, 2021 23:28:24 GMT
Results and discussion
Although the spike protein of SARS-CoV-2 can facilitate cell entry through well-known interactions between its RBD and ACE2,12 it is possible that the SARS-CoV-2 RBD may interact with other host molecules, including blood group antigens, which in turn may contribute to disease susceptibility. In particular, the RBD of SARS-CoV-2 contains general structural features similar to galectins, an ancient family of carbohydrate-binding proteins expressed in all metazoans. Accordingly, sequence alignment revealed sequence similarities between the SARS-CoV-2 RBD and human galectins (Figure 1A-B). Because galectins have been shown to exhibit high affinity for blood group antigens,15 we next examined whether the SARS-CoV-2 RBD may display similar blood group antigen recognition. To test this, we examined SARS-CoV-2 RBD binding with RBCs isolated from blood group A, B, or O individuals. Interestingly, the SARS-CoV-2 RBD exhibited only low-level binding to human RBCs of all types and failed to display any detectable preference for blood type A RBCs (Figure 1C; supplemental Figure 1). In contrast, the SARS-CoV-2 RBD readily bound ACE2-expressing HEK293 T cells, and anti-A antibody likewise bound blood group A RBCs (Figure 1D-E). Thus, significant binding of the SARS-CoV-2 RBD to the blood group A structures found on human blood group A RBCs does not seem to contribute to the increased likelihood of SARS-CoV-2 infection in blood group A individuals.
Figure 1.
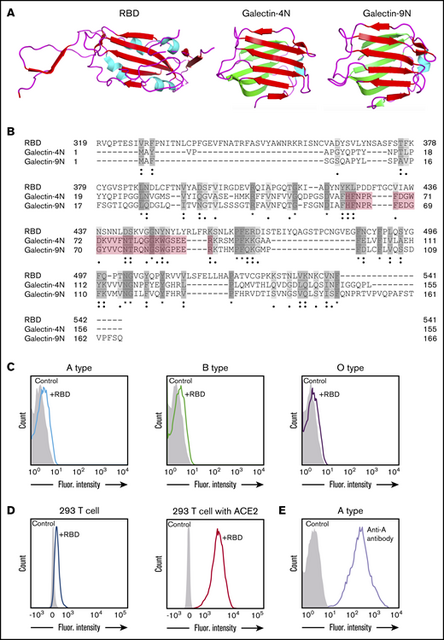
SARS-CoV-2 shares significant similarity to human galectins. (A) Structural representation of SARS-CoV-2 RBD (Protein Data Bank [PDB] ID: 6VXX), human galectin-4N-terminal domain (PDB: 5DUW), and human galectin-9N-terminal domain (PDB ID: 3LSD); β-sheet structures shown in red or green, α-helix shown in cyan. (B) Sequence alignment (Uniport align program) of SARS-CoV-2 RBD, human galectin-4N-terminal domain (galectin-4N) and human galectin-9 N-terminal domain (galectin-9N) indicating fully conserved residues (dark gray [*]), highly conserved residues (gray [:]), lowly conserved residues (light gray [.]); the galectin carbohydrate binding pocket is highlighted in light red. (C) Flow cytometric analysis of blood group A, B, or O type human RBCs after incubation with SARS-CoV-2 RBD for 1 hour followed by detection with anti-His antibody. (D) Flow cytometric analysis of HEK293 T cells alone or HEK293 T cells stably transduced to express ACE2 after a 1-hour incubation with the SARS-CoV-2 RBD and detection with anti-His antibody. (E) Flow cytometric examination of blood group A RBCs after incubation with anti-A antibody. Secondary antibody alone served as a control.
In addition to the ABO(H) blood group antigens expressed on RBCs, called type II ABO(H) blood group antigens, ABO(H) antigens are also expressed on various other tissues throughout the body including the respiratory epithelium.17,18 However, the types of ABO(H) antigens expressed along respiratory epithelium, referred to as type I ABO(H) antigens, are subtly, yet fundamentally different than their type II ABO(H) counterparts because of a distinct linkage configuration between the penultimate galactose residue and N-acetylglucosamine (type I, β1-3; type II, β1-4) (Figure 2A).19 Given its aerosolized route of infection, SARS-CoV-2 could have evolved a specific preference for the types of ABO(H) antigens expressed along respiratory epithelial cells. However, to specifically detect this possible preference, a completely distinct approach must be used. Because carbohydrates by definition are posttranslational modifications, they cannot easily be cloned and therefore rapidly expressed to explore these types of interactions. Instead, their synthesis often requires a complex approach of chemoenzymatic synthesis to generate distinct glycan libraries. To overcome this challenge, glycan microarrays populated with distinct glycan determinants have been generated to elucidate the fine specificity of carbohydrate-binding proteins for glycan determinants; such an approach has accurately predicted interactions with numerous cell types.20 As a result, we next turned to a glycan microarray format to define the possible specificity of SARS-CoV-2 for the distinct type I ABO(H) antigens expressed along the respiratory epithelium. In accordance with the lack of preferential blood group binding of the SARS-CoV-2 RBD to RBCs isolated from blood group A, B, or O individuals, no significant binding was observed toward the type II structures of A, B, or O(H) individuals (Figure 2B). In contrast, the SARS-CoV-2 RBD exhibited high preference for the same type of blood group A (type I) expressed on respiratory epithelial cells (Figure 2B). Earlier studies demonstrated that the association of ABO(H) blood group expression and infection is likewise shared by SARS-CoV.21 However, the underlying mechanism responsible for the increased propensity of blood group A individuals to become infected with SARS-CoV, similar to that in SARS-CoV-2, remained incompletely understood. Given the binding preference of SARS-CoV-2 RBD for type I blood group A, we next sought to determine whether a similar binding specificity existed for the SARS-CoV RBD. Despite sharing only 73% of its identity with SARS-CoV-2, SARS-CoV exhibited the same ABO(H) glycan-binding preference for the A antigen expressed in the respiratory tract (Figure 2C).
Figure 2.

SARS-CoV-2 RBD preferentially binds type I blood group A antigen. (A) Representation of human blood group antigens present on respiratory epithelium (type I) and RBCs (type II). Type I vs type II structures differ based on the linkage between the galactose and N-acetylglucosamine (type I: galactose β1-3 N-acetylglucosamine; type II: galactose β1-4 N-acetylglucosamine). (B-C) ABO(H) glycan microarray data obtained after incubation of SARS-CoV-2 RBD (B) or SARS-CoV RBD (C) with the corresponding glycans shown. Data are representative of 2 independent experiments. Error bars represent mean ± standard deviation. Statistics were generated by one-way analysis of variance with a post hoc Tukey’s multiple comparison. RFU, relative fluorescence unit. ***P < .001 for comparison between RBD binding to A type I vs all other glycans.
The capacity of the RBDs of SARS-CoV and SARS-CoV-2 that are responsible for initial host-cell contact to preferentially recognize the type of blood group A antigen uniquely expressed on respiratory epithelial cells may provide some insight into the apparent preference of SARS-CoV-2 and perhaps other severe corona viruses (SARS-CoV) for blood group A individuals. However, because these results do not definitively demonstrate that blood group A directly contributes to SARS-CoV-2 infection, future studies will certainly be needed to expand upon these initial findings, including an examination of the overall affinity and residues within the RBD responsible for blood group A interactions. Additional studies will likewise be needed to determine whether ABO(H) expression influences viral adhesion to different regions of the nasopharyngeal and respiratory tracts, actual infection of these cells in an ACE2-dependent or independent manner, or the overall stability of the virus along the mucosal surface. Furthermore, it should be noted that in addition to potentially influencing SARS-CoV-2 interactions with host cells, the impact of ABO(H) antigen expression on von Willebrand factor levels may likewise influence thromboembolic complications and therefore COVID-19 disease progression.22 Whatever the possible contribution of ABO(H) antigens to infection and possible disease progression, the ability of the SARS-CoV-2 to directly interact with the blood group A antigen uniquely expressed on respiratory epithelial cells provides clear evidence of a direct association between SARS-CoV-2 and the ABO(H) genetic locus.
Although the spike protein of SARS-CoV-2 can facilitate cell entry through well-known interactions between its RBD and ACE2,12 it is possible that the SARS-CoV-2 RBD may interact with other host molecules, including blood group antigens, which in turn may contribute to disease susceptibility. In particular, the RBD of SARS-CoV-2 contains general structural features similar to galectins, an ancient family of carbohydrate-binding proteins expressed in all metazoans. Accordingly, sequence alignment revealed sequence similarities between the SARS-CoV-2 RBD and human galectins (Figure 1A-B). Because galectins have been shown to exhibit high affinity for blood group antigens,15 we next examined whether the SARS-CoV-2 RBD may display similar blood group antigen recognition. To test this, we examined SARS-CoV-2 RBD binding with RBCs isolated from blood group A, B, or O individuals. Interestingly, the SARS-CoV-2 RBD exhibited only low-level binding to human RBCs of all types and failed to display any detectable preference for blood type A RBCs (Figure 1C; supplemental Figure 1). In contrast, the SARS-CoV-2 RBD readily bound ACE2-expressing HEK293 T cells, and anti-A antibody likewise bound blood group A RBCs (Figure 1D-E). Thus, significant binding of the SARS-CoV-2 RBD to the blood group A structures found on human blood group A RBCs does not seem to contribute to the increased likelihood of SARS-CoV-2 infection in blood group A individuals.
Figure 1.
SARS-CoV-2 shares significant similarity to human galectins. (A) Structural representation of SARS-CoV-2 RBD (Protein Data Bank [PDB] ID: 6VXX), human galectin-4N-terminal domain (PDB: 5DUW), and human galectin-9N-terminal domain (PDB ID: 3LSD); β-sheet structures shown in red or green, α-helix shown in cyan. (B) Sequence alignment (Uniport align program) of SARS-CoV-2 RBD, human galectin-4N-terminal domain (galectin-4N) and human galectin-9 N-terminal domain (galectin-9N) indicating fully conserved residues (dark gray [*]), highly conserved residues (gray [:]), lowly conserved residues (light gray [.]); the galectin carbohydrate binding pocket is highlighted in light red. (C) Flow cytometric analysis of blood group A, B, or O type human RBCs after incubation with SARS-CoV-2 RBD for 1 hour followed by detection with anti-His antibody. (D) Flow cytometric analysis of HEK293 T cells alone or HEK293 T cells stably transduced to express ACE2 after a 1-hour incubation with the SARS-CoV-2 RBD and detection with anti-His antibody. (E) Flow cytometric examination of blood group A RBCs after incubation with anti-A antibody. Secondary antibody alone served as a control.
In addition to the ABO(H) blood group antigens expressed on RBCs, called type II ABO(H) blood group antigens, ABO(H) antigens are also expressed on various other tissues throughout the body including the respiratory epithelium.17,18 However, the types of ABO(H) antigens expressed along respiratory epithelium, referred to as type I ABO(H) antigens, are subtly, yet fundamentally different than their type II ABO(H) counterparts because of a distinct linkage configuration between the penultimate galactose residue and N-acetylglucosamine (type I, β1-3; type II, β1-4) (Figure 2A).19 Given its aerosolized route of infection, SARS-CoV-2 could have evolved a specific preference for the types of ABO(H) antigens expressed along respiratory epithelial cells. However, to specifically detect this possible preference, a completely distinct approach must be used. Because carbohydrates by definition are posttranslational modifications, they cannot easily be cloned and therefore rapidly expressed to explore these types of interactions. Instead, their synthesis often requires a complex approach of chemoenzymatic synthesis to generate distinct glycan libraries. To overcome this challenge, glycan microarrays populated with distinct glycan determinants have been generated to elucidate the fine specificity of carbohydrate-binding proteins for glycan determinants; such an approach has accurately predicted interactions with numerous cell types.20 As a result, we next turned to a glycan microarray format to define the possible specificity of SARS-CoV-2 for the distinct type I ABO(H) antigens expressed along the respiratory epithelium. In accordance with the lack of preferential blood group binding of the SARS-CoV-2 RBD to RBCs isolated from blood group A, B, or O individuals, no significant binding was observed toward the type II structures of A, B, or O(H) individuals (Figure 2B). In contrast, the SARS-CoV-2 RBD exhibited high preference for the same type of blood group A (type I) expressed on respiratory epithelial cells (Figure 2B). Earlier studies demonstrated that the association of ABO(H) blood group expression and infection is likewise shared by SARS-CoV.21 However, the underlying mechanism responsible for the increased propensity of blood group A individuals to become infected with SARS-CoV, similar to that in SARS-CoV-2, remained incompletely understood. Given the binding preference of SARS-CoV-2 RBD for type I blood group A, we next sought to determine whether a similar binding specificity existed for the SARS-CoV RBD. Despite sharing only 73% of its identity with SARS-CoV-2, SARS-CoV exhibited the same ABO(H) glycan-binding preference for the A antigen expressed in the respiratory tract (Figure 2C).
Figure 2.
SARS-CoV-2 RBD preferentially binds type I blood group A antigen. (A) Representation of human blood group antigens present on respiratory epithelium (type I) and RBCs (type II). Type I vs type II structures differ based on the linkage between the galactose and N-acetylglucosamine (type I: galactose β1-3 N-acetylglucosamine; type II: galactose β1-4 N-acetylglucosamine). (B-C) ABO(H) glycan microarray data obtained after incubation of SARS-CoV-2 RBD (B) or SARS-CoV RBD (C) with the corresponding glycans shown. Data are representative of 2 independent experiments. Error bars represent mean ± standard deviation. Statistics were generated by one-way analysis of variance with a post hoc Tukey’s multiple comparison. RFU, relative fluorescence unit. ***P < .001 for comparison between RBD binding to A type I vs all other glycans.
The capacity of the RBDs of SARS-CoV and SARS-CoV-2 that are responsible for initial host-cell contact to preferentially recognize the type of blood group A antigen uniquely expressed on respiratory epithelial cells may provide some insight into the apparent preference of SARS-CoV-2 and perhaps other severe corona viruses (SARS-CoV) for blood group A individuals. However, because these results do not definitively demonstrate that blood group A directly contributes to SARS-CoV-2 infection, future studies will certainly be needed to expand upon these initial findings, including an examination of the overall affinity and residues within the RBD responsible for blood group A interactions. Additional studies will likewise be needed to determine whether ABO(H) expression influences viral adhesion to different regions of the nasopharyngeal and respiratory tracts, actual infection of these cells in an ACE2-dependent or independent manner, or the overall stability of the virus along the mucosal surface. Furthermore, it should be noted that in addition to potentially influencing SARS-CoV-2 interactions with host cells, the impact of ABO(H) antigen expression on von Willebrand factor levels may likewise influence thromboembolic complications and therefore COVID-19 disease progression.22 Whatever the possible contribution of ABO(H) antigens to infection and possible disease progression, the ability of the SARS-CoV-2 to directly interact with the blood group A antigen uniquely expressed on respiratory epithelial cells provides clear evidence of a direct association between SARS-CoV-2 and the ABO(H) genetic locus.