Post by Admin on Jan 2, 2022 2:13:16 GMT
Discussion
This study takes advantage of whole-genome sequence data from the 2500 years old Jomon individual, IK002, dissecting the origins of present-day East Asians. IK002 is modeled as a basal lineage to East Asians, Northeast Asians/East Siberians, and Native Americans (basal East Eurasians, bEE) after the divergence between Tiányuán and the ancestor of hunters-gatherers in Southeast Asia (Fig. 4)7,17. We clearly show the early divergence of IK002 from the common ancestor of the other ancient and present-day East Eurasian and Native Americans (Fig. 1c). Given that the split between the East Asian lineage and the Northeast Asians/East Siberian and Native American lineage was estimated to be 26 kya49, the divergence of the lineage leading to IK002 is likely to have occurred before this time but after 40 kya when the Tiányuán appeared (Fig. 4). Therefore, our results support the archaeological evidence based on lithic industry that the Jomon are direct descendants of the Upper-Paleolithic people who started living in the Japanese archipelago 38 kya (Supplementary Discussion).
Fig. 4: Schematic of peopling history in Southeast and East Asians, Northeast Asian/East Siberians and Native Americans.
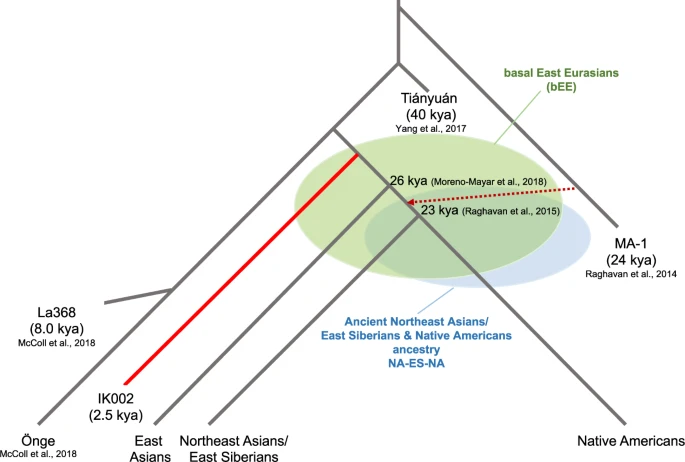
The basal East Eurasians (bEE) are an ancient population that had no divergence among the ancestors of East Asians, Northeast Asians/East Siberian, and Native Americans. NA-ES-NA presents another ancient population that had no split between the ancestors of Northeast Asians/East Siberian and Native Americans.
We use the MA-1 ancestry as a proxy for ancestral populations who took the northern route of Himalaya mountains to come to East Eurasia. The fine stone tool, i.e., microblade, is a representative technology that was originally developed around Lake Baikal in Central Siberia during the Upper-Paleolithic period9. This microblade culture was also reached the Hokkaido island ~25 kya and the main island of the Japanese archipelago ~20 kya. If this culture was brought by demic diffusion, IK002 should still retain the MA-1 ancestry. However, we find no evidence on the genetic affinity of MA-1 with ancient and present-day Southeast/East Asians including Devil′s Gate Cave (8.0 kya), Chokhopani (3.0–2.4 kya), and IK002 (2.5 kya) (Fig. 2). Therefore, we conclude that MA-1 gene flow occurred after the divergence between the ancestral populations of Northeast Asians/East Siberians/Native Americans (NA-ES-NA) and East Asians (Fig. 4): namely, East Asians originated in Southeast Asia without any detectable genetic influence from the ancestor who took the northern route. There are two hypothetical possibilities to explain the contradiction between genome data and archaeological records. The first possibility is that MA-1 may not be a direct ancestor who invented the microblade culture. The second is that, if the assumption is correct, then the northern-route culture represented by microblade was brought to the Japanese archipelago by the NA-ES-NA population who must have had substantial gene flow from MA-1, which was a typical “cultural diffusion.” The first and second possibilities can be examined by obtaining genome data of Upper-Paleolithic specimens hopefully accompanying with microblade newly excavated from around Lake Baikal and from the Primorye region, respectively.

The genome of the Ikawazu Jomon (IK002) strongly supports the dual structure model of the origins of modern Japanese37. The PCA plot and phylogenetic tree showed that the present-day Japanese fell in the cluster of present-day East Asians (e.g., Han Chinese) but not clustered with IK002 (Fig. 1a & c), while a signal of gene flow was detected from IK002 to present-day Japanese (Fig. 1c & Supplementary Fig. 8). The PCA and ADMIXTURE showed the close relationship between IK002 and the Hokkaido Ainu even in the genome-wide structure reflected by linkage blocks (Supplementary Fig. 4 & Supplementary Fig. 6). These results fit the hypothesis that the Ainu and the Jomon share the common ancestor: the present-day Honshu Japanese are the hybrid between the Jomon and migrants from the East Eurasian continent, and the Hokkaido Ainu have less influence of genetic contribution of the migrants37,42.
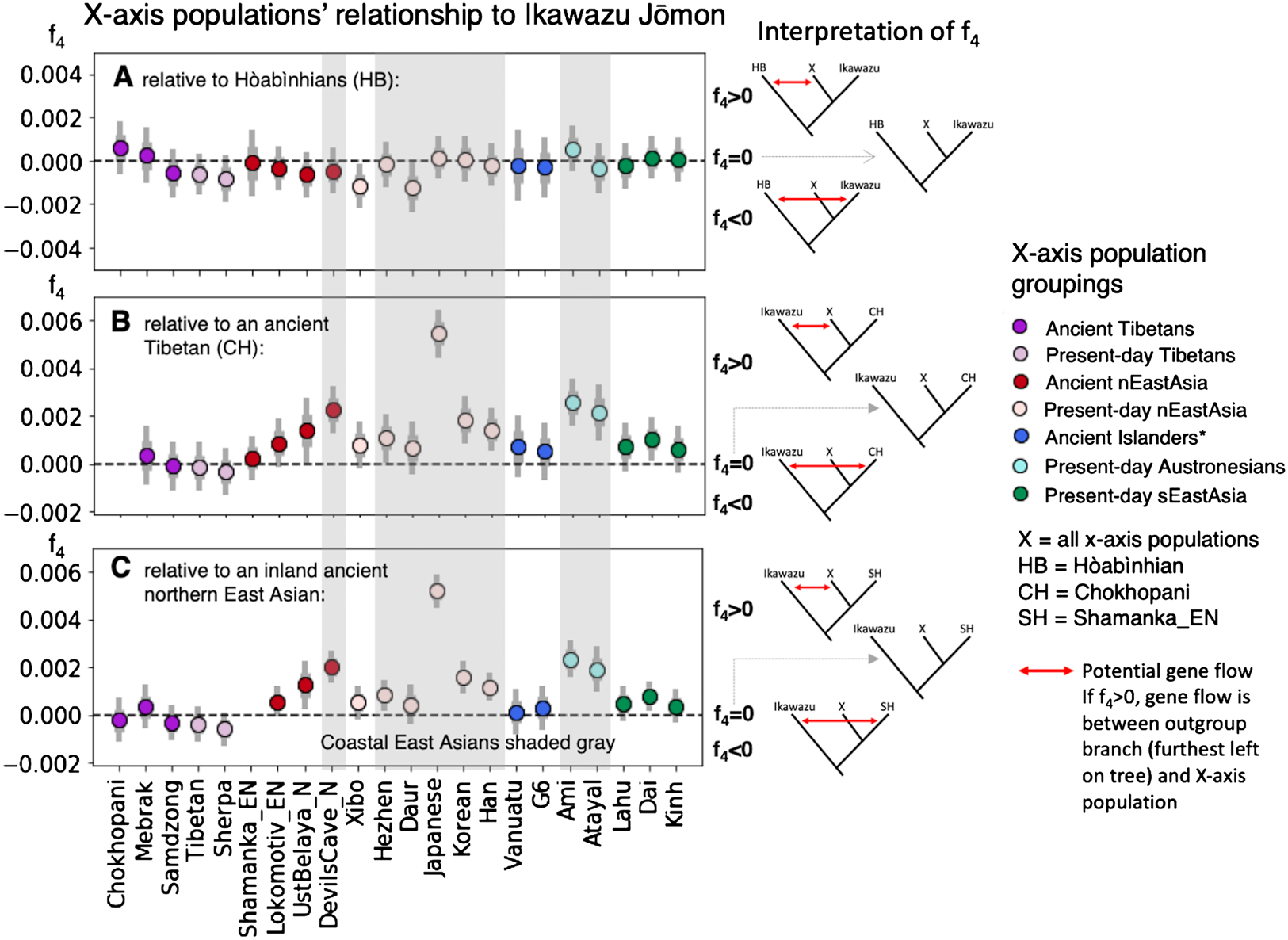
IK002 gave new insights into the migration route from south to north in East Eurasia. The f4-statistics suggest that both the ancient and the present-day East Asians are closer to IK002 than Chokhopani (ancient Tibetans, 3.0–2.4 kya) in the coastal region but not in the inland region (Fig. 3 & Supplementary Fig. 9). Recent archaeological evidence shows little cultural influence from ancestral Austronesians to the Ryukyu islands (i.e., southernmost islands in the Japanese archipelago) in the last 10,000 years51. Given this cultural continuity, it is unlikely that this extra genetic affinity to IK002 was formed due to recent gene flow as shown in a previous study52 (see Supplementary Discussion). Here, we provide two explanations for this signal: (1) the earliest-wave of migration from south to north occurred through the coastal region, and/or (2) the migration occurred in both the coastal and inland regions, but the genetic components of the earliest-wave were drowned out by back-migration(s) from north to south occurred in the inland region. In the early migration of anatomically modern humans, the route along the coast has been primarily thought to be important3,53,54,55,56. The use of water craft could support such explanation for the expansions through the islands and the coastal region3, which supports the first explanation. There could be, however, potential criticisms: such archaeological evidence of craft boat is hardly found. Ulchi and Nivkh show significantly negative values of f4-statistics (Z = −4.541 and −10.148, respectively. p < 0.000006). This could be an influence of the Hokkaido Ainu who are likely to be direct descendants of the Jomon people. The ancestor of the Ainu people could have admixed with the Okhotsk people57 who were genetically as well as morphologically close to Ulchi and Nivkh in the the Primorye region58,59,60,61,62. The second explanation is that the track of the earliest-wave was erased in the inland but left over in the coastal region. Taiwan aborigines (Ami and Atayal) and Igorot are the Austronesian minorities. Taiwan aborigines are thought to have come from the East Eurasian continent 13.2 ± 3.8 kya63, though the origin of Igorot (which can be derogatory, and could be best referred as Kankanaey) is not well-known.
The coastal-migration hypothesis would be supported by morphological traits of the Jomon individuals excavated from the Hobi shell-mound site, which is geographically very close to the Ikawazu shell-mound site in the Atsumi peninsula. Kaifu and Masuyama (2018) examined spatiotemporal variation in humeral shaft thickness using a sample of 1003 prehistoric individuals from various sites in the Japanese Archipelago, including 797 Jomon individuals64. The results showed that the Jomon humeri were thicker in coastal populations than in inland populations. Kaifu and Masuyama (2018) hypothesized that this was caused by not only fishing in the outer sea, but also active marine transportation by rowing boats. This previous study suggests the Hobi Jomon individuals in the Atsumi peninsula were the people who adapted to coastal environment.
However, there is still lack of ancient genome data in East Asia critical to understand the peopling history of East Eurasians. Although our data support the idea that IK002 was the direct descendant of the Upper-Paleolithic people, how/where those Upper-Paleolithic people migrated to the Japanese archipelago remains unanswered. It is required to analyze more ancient genome data in order to fill the gap and to prove the hypothetical migration routes.
Reviewers' comments:
Reviewer #1 (Remarks to the Author):
The manuscript focuses on genomic analysis of the 2.5 kya Jomon individual, IK002, an important piece of evidence that can inform us on the demographic history of eastern Eurasia. The authors concluded that IK002 is basal to all modern and ancient eastern Eurasians, and that, based the genetic affinity to Taiwanese aborigines, Jomon-related ancestry migrated into Japan from Southeast Asia likely via the coastal route.
2) Paragraph 2, Line 12 of Introduction: ‘This direct evidence on the link between the Jomon and Southeast Asians, thus, confirms the southern route origin of East Asians.’ The evidence only shows genetic affinity between Jomon and Hoabinhian-related ancestry. Most ancient and present-day Southeast Asians and East Asians do not show strong genetic affiliations with Hoabinhian individuals. The finding can only support southern route origin of Jomon individual, and not all of East Asians. The populations of Southeast Asia and East Asia are already structured since the Paleolithic (Hoabinhian-related, Siberian-related, and ancestors of Austronesian-related and Austroasiatic-related populations), and some of this may have migrated from north of the Himalayan arc.
7) Paragraph 5, Line1 or Resutls. The basal position of IK002 relative to the selected East Asian and Native American groups can be attributed to genetic affiliation of IK002 to Hoabinhian-related ancestry, which was not accounted for in the Treemix run. Figure S38 of McColl et al., 2019 models IK002 as an admixture between Hoabinhian-related and East Asian related ancestry (represented by Ami). How do you reconcile the difference between the result of this treemix run and the earlier published qpGraph analysis? Would the topology be consistent if you include other present-day Southeast Asians, Siberian (Koryak, Yakut, etc), and Papuans in the Treemix analysis?
api.repository.cam.ac.uk/server/api/core/bitstreams/0683d8fc-8266-40a9-bdc9-49242e86caef/content
This study takes advantage of whole-genome sequence data from the 2500 years old Jomon individual, IK002, dissecting the origins of present-day East Asians. IK002 is modeled as a basal lineage to East Asians, Northeast Asians/East Siberians, and Native Americans (basal East Eurasians, bEE) after the divergence between Tiányuán and the ancestor of hunters-gatherers in Southeast Asia (Fig. 4)7,17. We clearly show the early divergence of IK002 from the common ancestor of the other ancient and present-day East Eurasian and Native Americans (Fig. 1c). Given that the split between the East Asian lineage and the Northeast Asians/East Siberian and Native American lineage was estimated to be 26 kya49, the divergence of the lineage leading to IK002 is likely to have occurred before this time but after 40 kya when the Tiányuán appeared (Fig. 4). Therefore, our results support the archaeological evidence based on lithic industry that the Jomon are direct descendants of the Upper-Paleolithic people who started living in the Japanese archipelago 38 kya (Supplementary Discussion).
Fig. 4: Schematic of peopling history in Southeast and East Asians, Northeast Asian/East Siberians and Native Americans.
The basal East Eurasians (bEE) are an ancient population that had no divergence among the ancestors of East Asians, Northeast Asians/East Siberian, and Native Americans. NA-ES-NA presents another ancient population that had no split between the ancestors of Northeast Asians/East Siberian and Native Americans.
We use the MA-1 ancestry as a proxy for ancestral populations who took the northern route of Himalaya mountains to come to East Eurasia. The fine stone tool, i.e., microblade, is a representative technology that was originally developed around Lake Baikal in Central Siberia during the Upper-Paleolithic period9. This microblade culture was also reached the Hokkaido island ~25 kya and the main island of the Japanese archipelago ~20 kya. If this culture was brought by demic diffusion, IK002 should still retain the MA-1 ancestry. However, we find no evidence on the genetic affinity of MA-1 with ancient and present-day Southeast/East Asians including Devil′s Gate Cave (8.0 kya), Chokhopani (3.0–2.4 kya), and IK002 (2.5 kya) (Fig. 2). Therefore, we conclude that MA-1 gene flow occurred after the divergence between the ancestral populations of Northeast Asians/East Siberians/Native Americans (NA-ES-NA) and East Asians (Fig. 4): namely, East Asians originated in Southeast Asia without any detectable genetic influence from the ancestor who took the northern route. There are two hypothetical possibilities to explain the contradiction between genome data and archaeological records. The first possibility is that MA-1 may not be a direct ancestor who invented the microblade culture. The second is that, if the assumption is correct, then the northern-route culture represented by microblade was brought to the Japanese archipelago by the NA-ES-NA population who must have had substantial gene flow from MA-1, which was a typical “cultural diffusion.” The first and second possibilities can be examined by obtaining genome data of Upper-Paleolithic specimens hopefully accompanying with microblade newly excavated from around Lake Baikal and from the Primorye region, respectively.
The genome of the Ikawazu Jomon (IK002) strongly supports the dual structure model of the origins of modern Japanese37. The PCA plot and phylogenetic tree showed that the present-day Japanese fell in the cluster of present-day East Asians (e.g., Han Chinese) but not clustered with IK002 (Fig. 1a & c), while a signal of gene flow was detected from IK002 to present-day Japanese (Fig. 1c & Supplementary Fig. 8). The PCA and ADMIXTURE showed the close relationship between IK002 and the Hokkaido Ainu even in the genome-wide structure reflected by linkage blocks (Supplementary Fig. 4 & Supplementary Fig. 6). These results fit the hypothesis that the Ainu and the Jomon share the common ancestor: the present-day Honshu Japanese are the hybrid between the Jomon and migrants from the East Eurasian continent, and the Hokkaido Ainu have less influence of genetic contribution of the migrants37,42.
IK002 gave new insights into the migration route from south to north in East Eurasia. The f4-statistics suggest that both the ancient and the present-day East Asians are closer to IK002 than Chokhopani (ancient Tibetans, 3.0–2.4 kya) in the coastal region but not in the inland region (Fig. 3 & Supplementary Fig. 9). Recent archaeological evidence shows little cultural influence from ancestral Austronesians to the Ryukyu islands (i.e., southernmost islands in the Japanese archipelago) in the last 10,000 years51. Given this cultural continuity, it is unlikely that this extra genetic affinity to IK002 was formed due to recent gene flow as shown in a previous study52 (see Supplementary Discussion). Here, we provide two explanations for this signal: (1) the earliest-wave of migration from south to north occurred through the coastal region, and/or (2) the migration occurred in both the coastal and inland regions, but the genetic components of the earliest-wave were drowned out by back-migration(s) from north to south occurred in the inland region. In the early migration of anatomically modern humans, the route along the coast has been primarily thought to be important3,53,54,55,56. The use of water craft could support such explanation for the expansions through the islands and the coastal region3, which supports the first explanation. There could be, however, potential criticisms: such archaeological evidence of craft boat is hardly found. Ulchi and Nivkh show significantly negative values of f4-statistics (Z = −4.541 and −10.148, respectively. p < 0.000006). This could be an influence of the Hokkaido Ainu who are likely to be direct descendants of the Jomon people. The ancestor of the Ainu people could have admixed with the Okhotsk people57 who were genetically as well as morphologically close to Ulchi and Nivkh in the the Primorye region58,59,60,61,62. The second explanation is that the track of the earliest-wave was erased in the inland but left over in the coastal region. Taiwan aborigines (Ami and Atayal) and Igorot are the Austronesian minorities. Taiwan aborigines are thought to have come from the East Eurasian continent 13.2 ± 3.8 kya63, though the origin of Igorot (which can be derogatory, and could be best referred as Kankanaey) is not well-known.
The coastal-migration hypothesis would be supported by morphological traits of the Jomon individuals excavated from the Hobi shell-mound site, which is geographically very close to the Ikawazu shell-mound site in the Atsumi peninsula. Kaifu and Masuyama (2018) examined spatiotemporal variation in humeral shaft thickness using a sample of 1003 prehistoric individuals from various sites in the Japanese Archipelago, including 797 Jomon individuals64. The results showed that the Jomon humeri were thicker in coastal populations than in inland populations. Kaifu and Masuyama (2018) hypothesized that this was caused by not only fishing in the outer sea, but also active marine transportation by rowing boats. This previous study suggests the Hobi Jomon individuals in the Atsumi peninsula were the people who adapted to coastal environment.
However, there is still lack of ancient genome data in East Asia critical to understand the peopling history of East Eurasians. Although our data support the idea that IK002 was the direct descendant of the Upper-Paleolithic people, how/where those Upper-Paleolithic people migrated to the Japanese archipelago remains unanswered. It is required to analyze more ancient genome data in order to fill the gap and to prove the hypothetical migration routes.
Reviewers' comments:
Reviewer #1 (Remarks to the Author):
The manuscript focuses on genomic analysis of the 2.5 kya Jomon individual, IK002, an important piece of evidence that can inform us on the demographic history of eastern Eurasia. The authors concluded that IK002 is basal to all modern and ancient eastern Eurasians, and that, based the genetic affinity to Taiwanese aborigines, Jomon-related ancestry migrated into Japan from Southeast Asia likely via the coastal route.
2) Paragraph 2, Line 12 of Introduction: ‘This direct evidence on the link between the Jomon and Southeast Asians, thus, confirms the southern route origin of East Asians.’ The evidence only shows genetic affinity between Jomon and Hoabinhian-related ancestry. Most ancient and present-day Southeast Asians and East Asians do not show strong genetic affiliations with Hoabinhian individuals. The finding can only support southern route origin of Jomon individual, and not all of East Asians. The populations of Southeast Asia and East Asia are already structured since the Paleolithic (Hoabinhian-related, Siberian-related, and ancestors of Austronesian-related and Austroasiatic-related populations), and some of this may have migrated from north of the Himalayan arc.
7) Paragraph 5, Line1 or Resutls. The basal position of IK002 relative to the selected East Asian and Native American groups can be attributed to genetic affiliation of IK002 to Hoabinhian-related ancestry, which was not accounted for in the Treemix run. Figure S38 of McColl et al., 2019 models IK002 as an admixture between Hoabinhian-related and East Asian related ancestry (represented by Ami). How do you reconcile the difference between the result of this treemix run and the earlier published qpGraph analysis? Would the topology be consistent if you include other present-day Southeast Asians, Siberian (Koryak, Yakut, etc), and Papuans in the Treemix analysis?
api.repository.cam.ac.uk/server/api/core/bitstreams/0683d8fc-8266-40a9-bdc9-49242e86caef/content