|
|
Post by Admin on Jul 27, 2023 17:59:33 GMT
Extended Data Fig. 1: Tell settlements and burial sites in Southeastern Europe. a, Aerial view of Tell Măgura Gorgana near Pietrele, Romania (© Konstantin Scheele, German Archaeological Institute, Eurasia Department). b, Detailed view of the 11m stratigraphy at Pietrele (© Svend Hansen, German Archaeological Institute, Eurasia Department). c, Aerial view of Tell Yunatsite, Bulgaria (© Kamen Boyadzhiev). d, Map of the site Orlovka-Kartal, Ukraine. The base map was sourced from Google Earth www.google.com/earth/index.html. e, Characteristic finds from the Eneolithic type-site Usatove. f, Characteristic finds attributed to the Cernavoda I phase. g, Burial in flexed position from grave 10 at Kartal (© Igor Bruyako). h, Infant urn-burial from the Early Bronze Age layer south of tell Yunatsite (© Kamen Boyadzhiev).  Extended Data Fig. 2 F4-statistics for CA groups to determine Neolithic proxies. F4-statistics show different attraction of the CA to Neolithic groups conditioned on HG groups. Z-scores outside the threshold of (|Z|≥1) are highlighted in orange, f4-values are shown with one standard error. Test populations are given on the y-axis. Standard errors (SE) were computed with the default block jackknife approach. 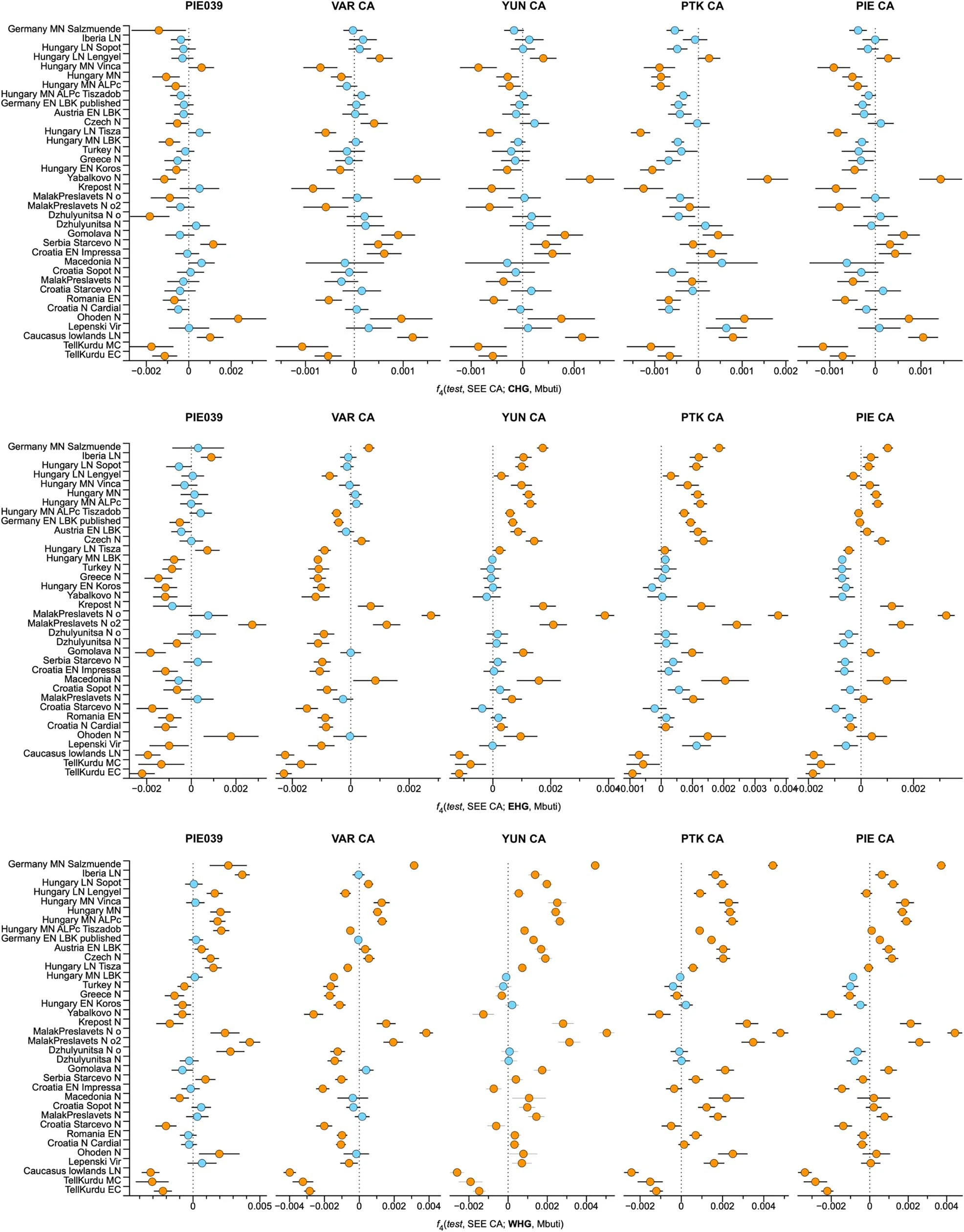 |
|
|
|
Post by Admin on Jul 28, 2023 19:08:28 GMT
Extended Data Fig. 3 DATES and Y-chromosomal haplogroup diversity. a, DATES decay curve for the CA outlier individual PIE060 with SEE N and Iron Gates HGs as source populations. b, Changes of Y-chromosomal haplogroup diversity (colour fills) over time (mean 14C dates cal. BC; x-axis) with respect to changes in autosomal ancestry as reflected in PC2 (y-axis), based on the relative density of female (open circles) and male (colour filled squares) of all newly reported individuals in this study. 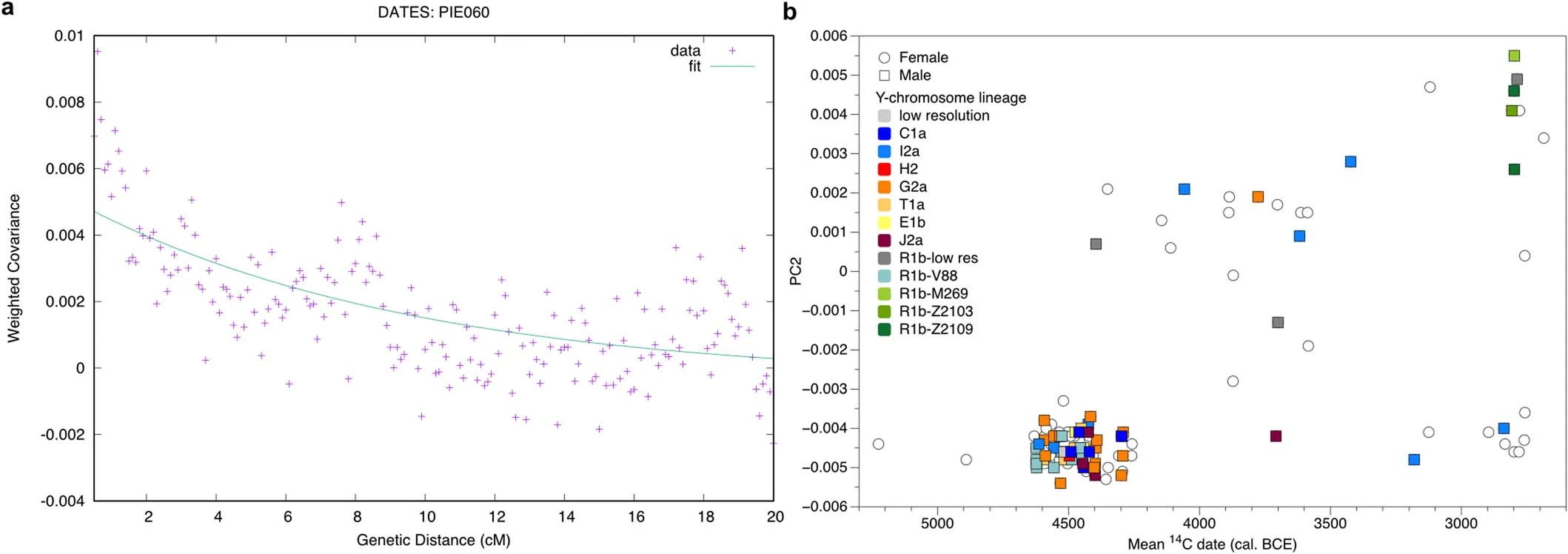 Extended Data Fig. 4 Identity-by-descent within and between sites. a, Results of identity-by-descent (IBD) analysis per pair of individuals. Plotting the sum versus the number of the shared chunks of IBD in window sizes of >12cM resolves degrees of biological relatedness up to the 4-6th degree. Within (left) and between site (right) relationships are highlighted separately. b, Stacked bar plot showing the number (left) and the cumulative distribution of the sum (right) of IBD blocks that are shared between all individuals within and between sites. 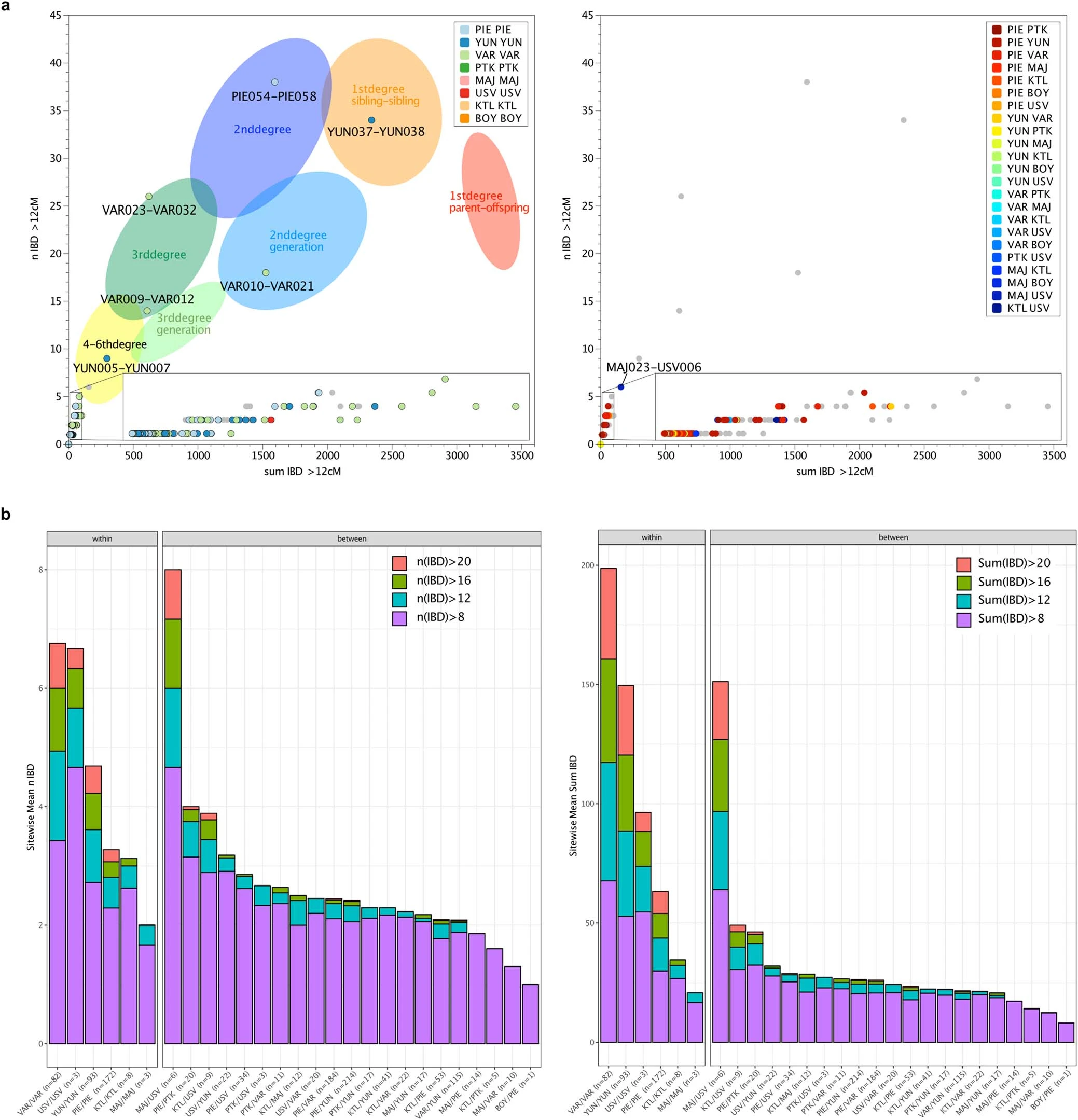 |
|
|
|
Post by Admin on Jul 29, 2023 17:58:18 GMT
Extended Data Fig. 5 Cumulative distribution of the runs of homozygosity tracts of all newly reported individuals. Runs of homozygosity were estimated with hapROH. Individuals are grouped in relative chronological order from right to left. Expected parental relationship and simulated effective populations sizes are given. 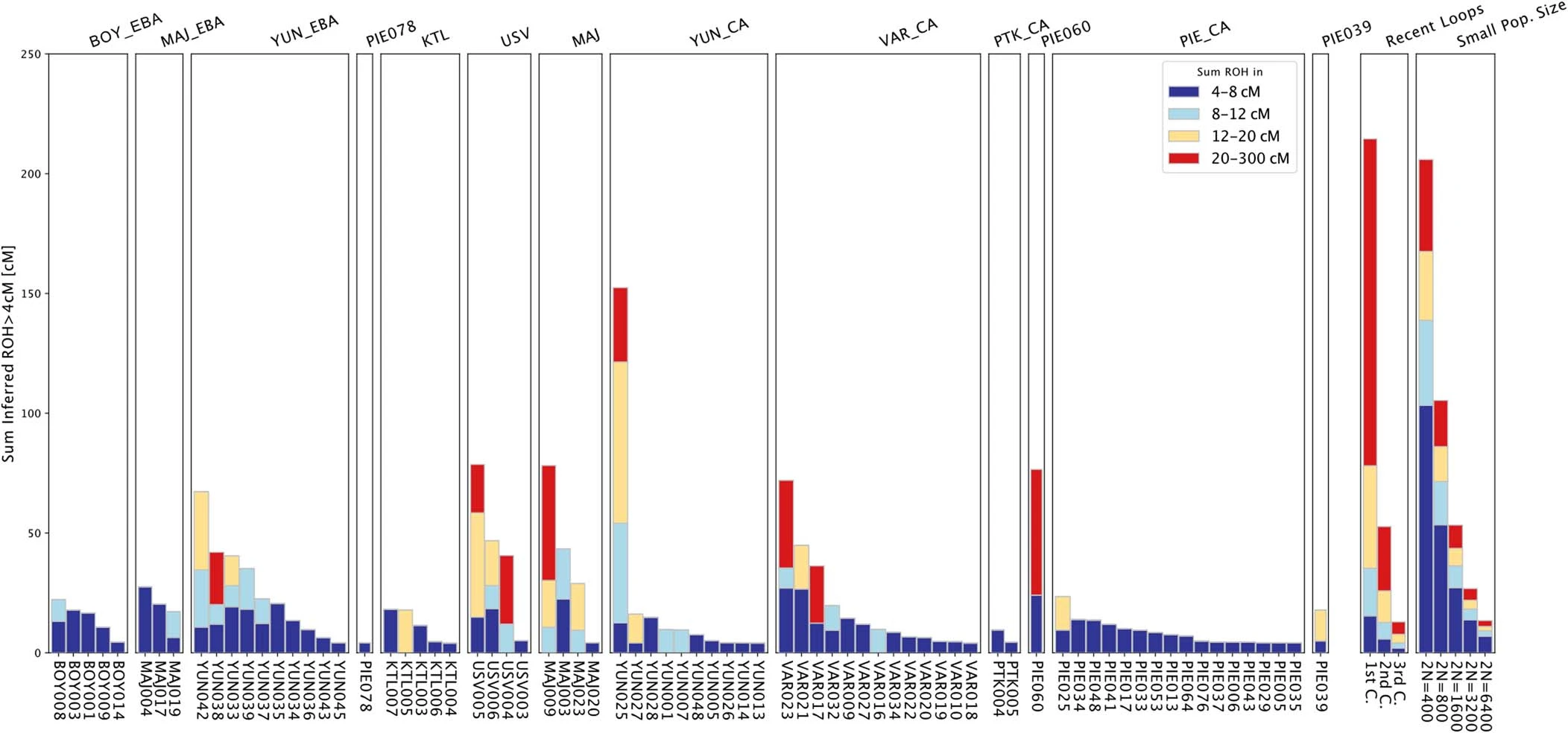 Extended Data Fig. 6 F4 statistics for Ukraine Eneolithic groups to determine attraction to ‘cornerstone’ populations. F4 statistics show different attractions of Ukraine Eneolithic groups to ‘cornerstone’ ancestry groups conditioned on a, Maykop-associated groups, b, Steppe Eneolithic, and c, SEE CA. Significant Z-scores (|Z|≥3) are highlighted in orange, f4 values are shown with three standard errors. Test populations are given on the y-axis. Standard errors (SE) were computed with the default block jackknife approach. 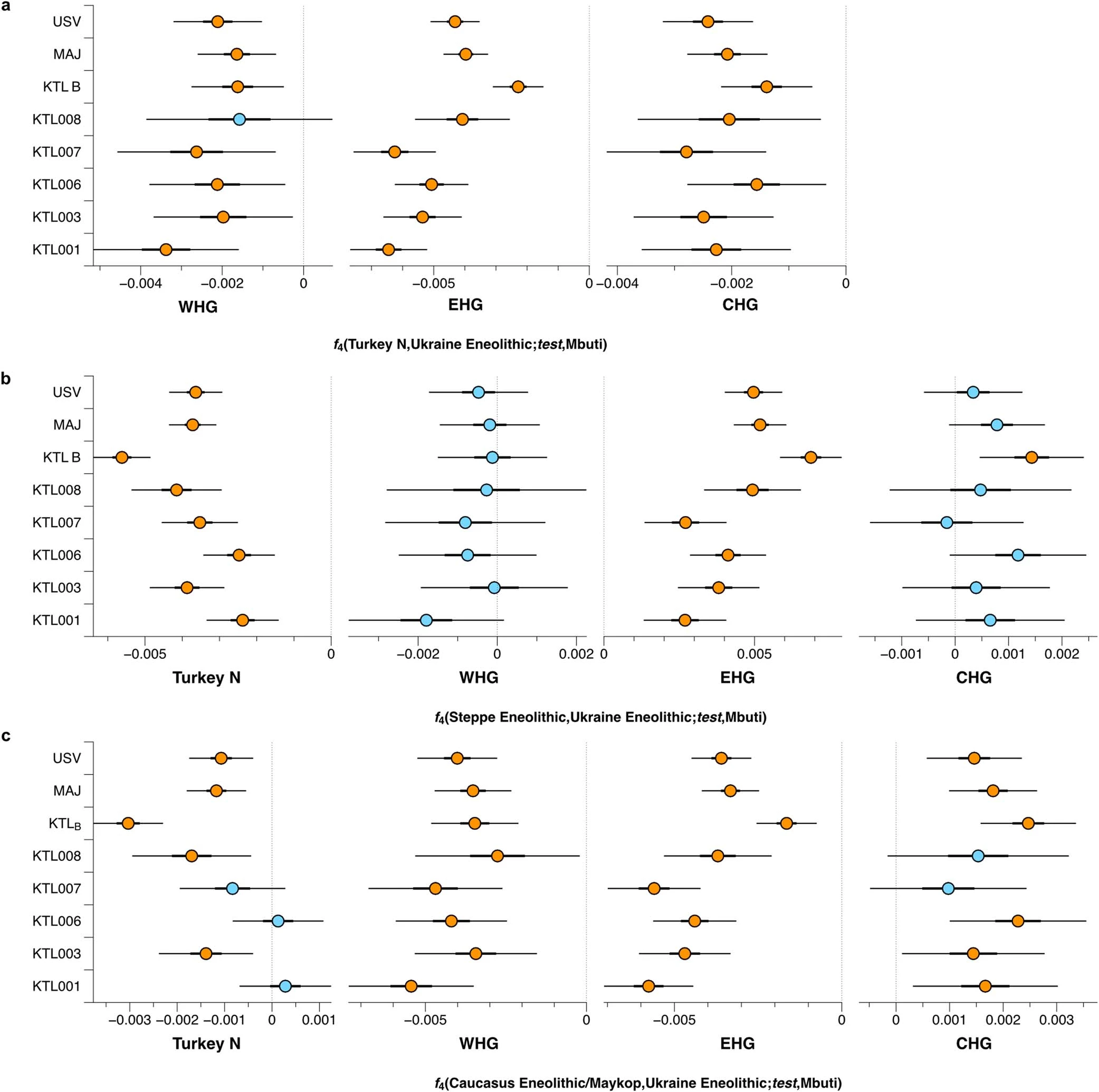 |
|
|
|
Post by Admin on Jul 30, 2023 18:05:35 GMT
Extended Data Fig. 7 F4 statistics for EBA groups to test for excess HG attraction. F4 statistics show different attractions of EBA groups to HG groups conditioned on their respective preceding or contemporaneous group. Significant Z-scores (|Z|≥3) are highlighted in orange, f4 values are shown with three standard errors. Test populations are given on the y-axis. Standard errors (SE) were computed with the default block jackknife approach. 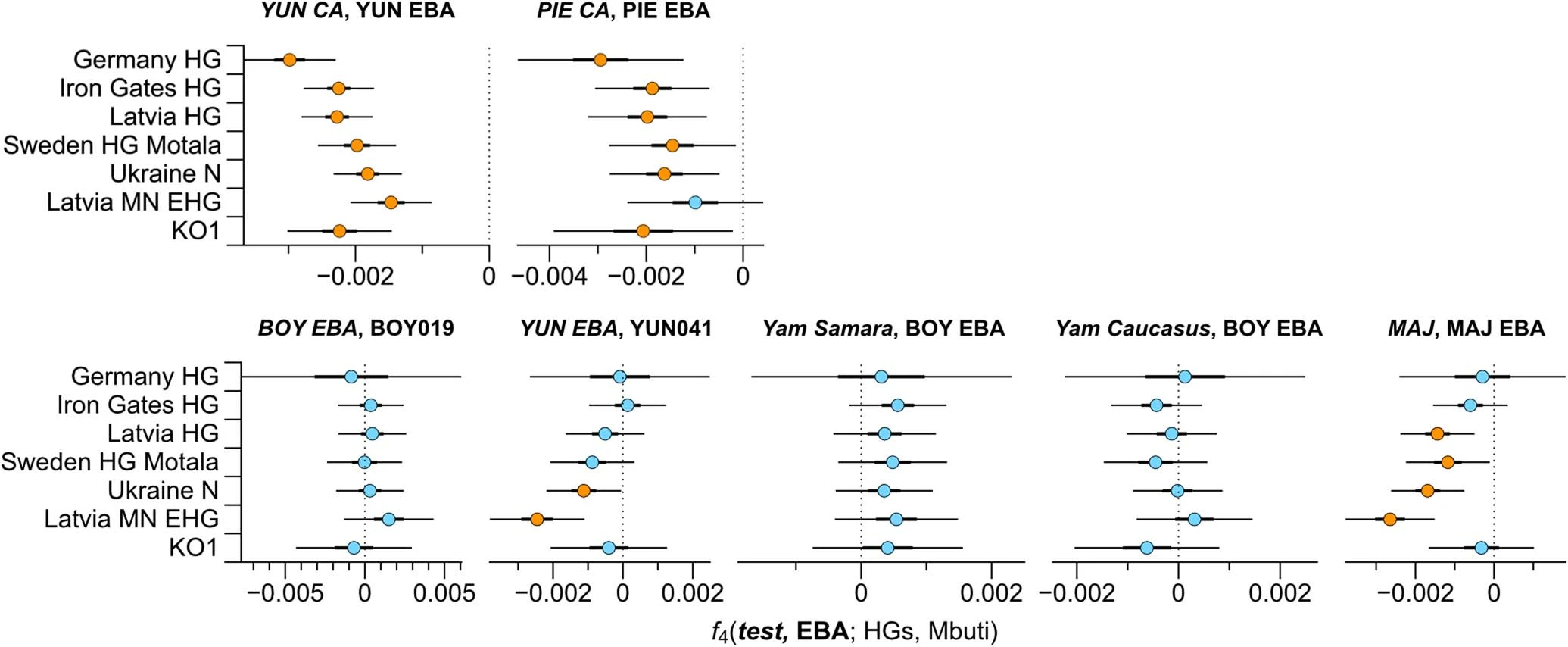 Extended Data Fig. 8 Testing for Anatolian farmer-related affinity in EBA individuals. F4 statistics show different attractions of the EBA groups to VAR_CA conditioned on Yamnaya-associated groups. Significant Z-scores (|Z|≥3) are highlighted in orange, f4 values are shown with three standard errors. Test populations are given on the y-axis. Standard errors (SE) were computed with the default block jackknife approach. 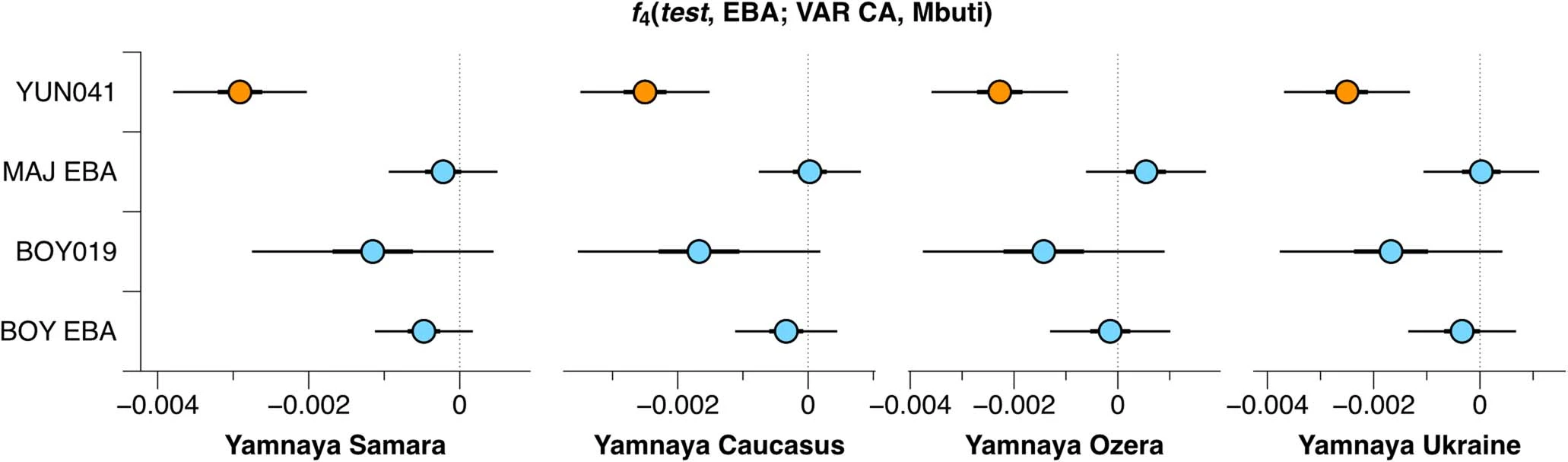 Extended Data Fig. 9 F4 statistics to explore excess affinity of EBA ‘steppe ancestry’ groups conditioned on preceding Eneolithic groups from the steppe and the Caucasus. F4 statistics show different attractions of the EBA groups to ‘cornerstone’ populations conditioned on pre-Yamnaya groups, Steppe Eneolithic and Caucasus Eneolithic/Maykop. Significant Z-scores (|Z|≥3) are highlighted in orange, f4 values are shown with three standard errors. EBA groups are given on the y-axis and test populations are given on the x-axis. Standard errors (SE) were computed with the default block jackknife approach. 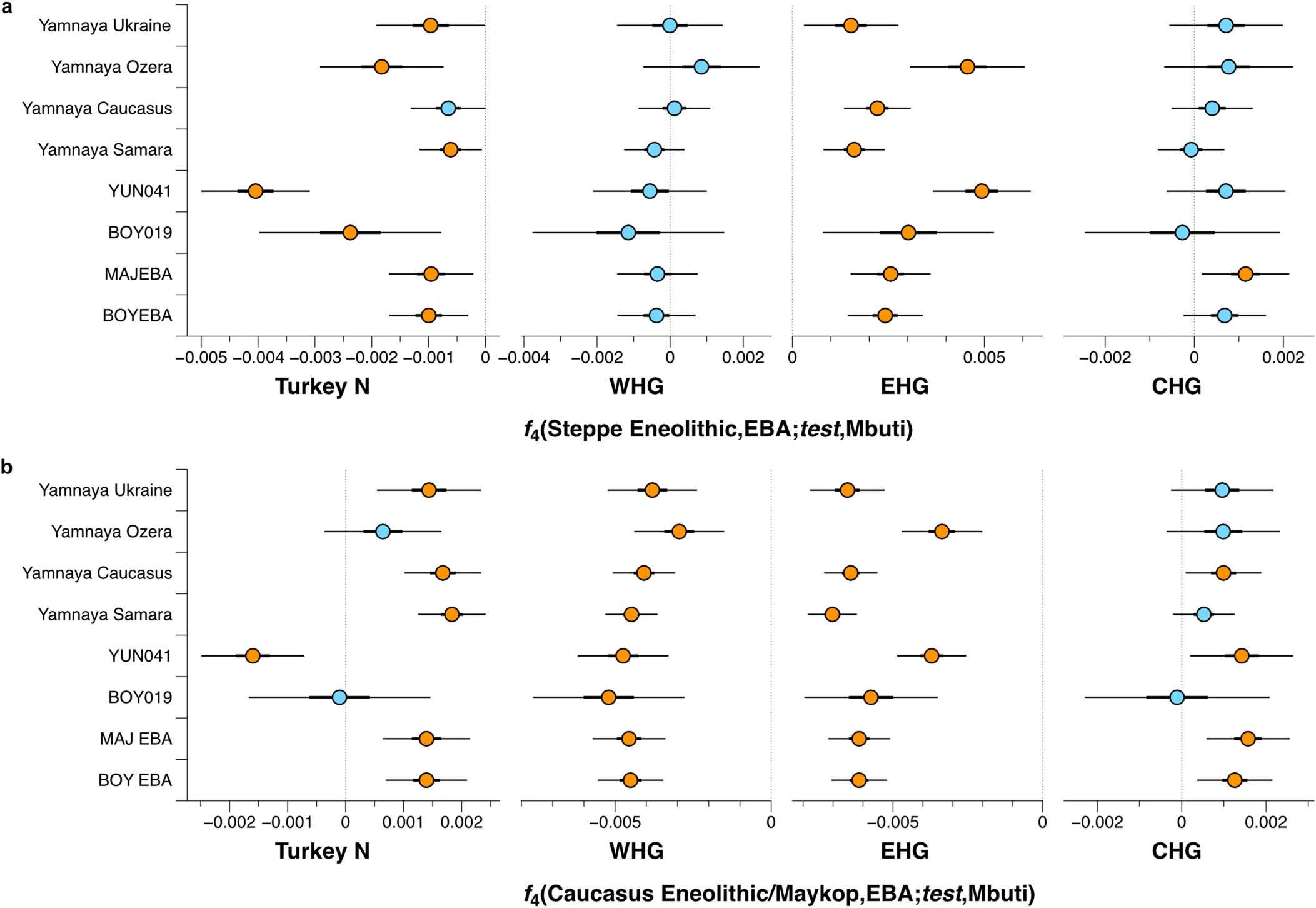 Early contact between late farming and pastoralist societies in southeastern Europe 1. Southeastern Europe 1.1 Ecological background
During the Late Neolithic and the Copper Age, climatically optimal conditions
prevailed. A continental climate with cold dry winters and moist early summers must have been
the norm, which is reflected in the adaptation and choice of cultivated and exploited plants. The
main attested species in the macro-botanical assemblage are einkorn and emmer wheat, barley,
lentils, peas and bitter vetch. Collected fruits included raspberry, cornelian cherry, cherry plum, grapes and blackthorn.
The paleoenvironmental studies at tell Pietrele have provided evidence that the Danube did not
exist in the form of a streamlined river as we know it today13. In addition to many side-arms,
the Danube to the east of the city of Ruse was rather a lake that at times probably reached as
far as the Black Sea.
This meant an almost inexhaustible supply of fish and mollusks for the inhabitants at
this lake. In Pietrele for example, mainly carp and pike were fished, and to a lesser extent also
catfish. While the older layers of this tell are mainly characterized by domesticated animals,
especially pigs, but also cattle and sheep, in the younger layers half of the animals are wild
game, indicating that the hunting of wild boar, red deer, wild horse and aurochs played a
substantial role95. Comparable results of the palaeobotanical and palaeozoological
investigations are also available from other tell settlements.
The end of the tell settlements has been linked, above all by Henrieta Todorova, to a
rise in annual mean temperature that led to long-lasting droughts and forest fires and finally to
a prolonged depopulation over several centuries16.
|
|
|
|
Post by Admin on Jun 27, 2024 17:23:57 GMT
The Genetic History of the South Caucasus from the Bronze to the Early Middle Ages: 5000 years of genetic continuity despite high mobility Abstract Archaeological and archaeogenetic studies have highlighted the pivotal role of the Caucasus region throughout prehistory, serving as a central hub for cultural, technological, and linguistic innovations. However, despite its dynamic history, the critical area between the Greater and Lesser Caucasus mountain ranges, mainly corresponding to modern-day Georgia, has received limited attention. Here, we generated an ancient DNA time transect consisting of 219 individuals with genome-wide data from 47 sites in this region, supplemented by 97 new radiocarbon dates. Spanning from the Early Bronze Age 5000 years ago to the so-called ‘Migration Period’ that followed the fall of the Western Roman Empire, we document a largely persisting local gene pool that continuously assimilated migrants from Anatolia/Levant and the populations of the adjacent Eurasian steppe. More specifically, we observe these admixture events as early as the Middle Bronze Age. Starting with Late Antiquity (late first century AD), we also detect an increasing number of individuals with more southern ancestry, more frequently associated with urban centers – landmarks of the early Christianization in eastern Georgia. Finally, in the Early Medieval Period starting 400 AD, we observe genetic outlier individuals with ancestry from the Central Eurasian steppe, with artificial cranial deformations (ACD) in several cases. At the same time, we reveal that many individuals with ACD descended from native South Caucasus groups, indicating that the local population likely adopted this cultural practice. Introduction Situated at the crossroads of Eastern Europe and Southwest Asia, the South Caucasus has long been recognized as a seminal center of cultural and technological innovation throughout prehistory 1–4. Despite being surrounded by formidable geographical barriers, such as the Greater Caucasus Mountain Range, the area constituted an isthmus connecting various civilizations 1–3. As early as the Bronze Age (BA; starting c. mid-4th millennium BC), the distinct traditions of the cultural complexes of Maykop and Kura-Araxes suggest a remarkable degree of cultural connectivity across the area south and north of this mountain range 4,5. Previous ancient DNA (aDNA) studies have revealed that the Caucasus formed no lasting barrier to human movement. The populations of the North Caucasus inhabiting the so-called piedmont region were genetically connected to the south since the Eneolithic 6, while the prehistoric populations of the South Caucasus and Anatolia extensively mixed since the 7th millennium BC 7. Conversely, the impact of mobility from BA steppe pastoralists into the South Caucasus has been attested using aDNA from the Middle/Late BA (c. 1900-1200 BC) onwards in Armenia 8- and also in eight genomes from two Late BA (LBA) sites in Georgia 9. Right after the formation of stratified societies in the Middle BA, the rise of political authorities in the LBA and Early Iron Age has been postulated 10,11. Beginning around 800 BC and coinciding with the linguistic diversification of proto-Karto/Georgian-Zan (a distinct language family from Indo-European) into Georgian and Zan12, the social and economic division of the region of present-day Georgia deepened, culminating in the formation of the first states of Colchis in the Iron Age, and the Kingdom of Iberia in Early Antiquity, respectively 12. Soon, Colchis was involved in the conflict against the Urartian and subsequent Median empires and finally fell under the rule of the Persian Achaemenid Empire 13. No later than the 6th century BC, foreign contacts through the Greek colonies were established on the Black Sea coast 14. On the eastern side of Georgia, the kingdom of Iberia was founded in the aftermath of Alexander’s conquest and, together with Colchis, were under the suzerainty of the Seleucid empire. Around 65 BC, the Romans conquered Colchis from the Kingdom of Pontus 15 and the Kingdom of Iberia, exposing it to the Graeco-Roman world and ultimately claiming both regions under the principate 14. The early Christianization of Iberia started in the 4th century AD 13 and remained strong considering the fluctuating political boundaries elsewhere on Rome’s eastern frontier 16. Since the 3rd century, initially Romans and Sassanians, and later Byzantines and Arabs, vehemently competed for Iberia and Colchis (later called Lazica) until the great medieval Kingdom of Georgia was unified in 1008 17. The South Caucasus has been a melting pot of diverse ethnic groups from as far away as Inner Asia and Eastern Europe, including Sarmatians and the Alans 18,19 and later the Huns during the migration period of the Early Middle Ages 20,21. These nomadic people of the steppes practiced certain types of artificial cranial deformations (ACD) 22. This practice also appears in the region of Georgia, evidencing influence from the north across the Caucasus in different periods 39,40. It remains unclear how the dynamic history of the South Caucasus region drove human mobility at the individual and group levels. Here, we generated an ancient genome-wide dataset of 219 individuals from 47 sites in Georgia. Importantly, we also produced 97 same-sample radiocarbon dates that confirm that sampling spans c. 4,500 years continuously from the EBA to the Early-High Middle Ages. This extensive time transect fills several critical gaps in archaeogenetic sampling for the area between the Greater and Lesser Caucasus, thus far represented by only eight Bronze Age 9 and two Upper Palaeolithic genomes (‘Caucasus hunter-gatherers’) 23. This new data enabled us to trace individual and group-level ancestry patterns through time. Our findings reveal a high degree of genetic continuity, along with lasting mixture events since the MBA with steppe-related and Anatolian/Levantine groups. From the early Christianization of the Iberia kingdom, we observe a high rate of individuals with ancestry from Anatolia or the Levant. Notably, we find that most of the individuals with ACD descended from native South Caucasus groups, highlighting a cultural practice adopted by the local population. Finally, by screening for identical-by-descent (IBD) segments, we establish higher levels of biological relatedness among individuals from rural areas buried a few kilometers apart, compared to large urban centers like the ancient capital of the Iberia kingdom. By unraveling the intricate tapestry of population movements and cultural exchanges, our results contribute to a better understanding of the region’s rich and diverse heritage. www.biorxiv.org/content/10.1101/2024.06.11.597880v1.full |
|

