|
|
Post by Admin on Jul 22, 2023 19:54:00 GMT
Early contact between late farming and pastoralist societies in southeastern Europe Abstract Archaeogenetic studies have described two main genetic turnover events in prehistoric western Eurasia: one associated with the spread of farming and a sedentary lifestyle starting around 7000–6000 BC (refs. 1,2,3) and a second with the expansion of pastoralist groups from the Eurasian steppes starting around 3300 BC (refs. 4,5). The period between these events saw new economies emerging on the basis of key innovations, including metallurgy, wheel and wagon and horse domestication6,7,8,9. However, what happened between the demise of the Copper Age settlements around 4250 BC and the expansion of pastoralists remains poorly understood. To address this question, we analysed genome-wide data from 135 ancient individuals from the contact zone between southeastern Europe and the northwestern Black Sea region spanning this critical time period. While we observe genetic continuity between Neolithic and Copper Age groups from major sites in the same region, from around 4500 BC on, groups from the northwestern Black Sea region carried varying amounts of mixed ancestries derived from Copper Age groups and those from the forest/steppe zones, indicating genetic and cultural contact over a period of around 1,000 years earlier than anticipated. We propose that the transfer of critical innovations between farmers and transitional foragers/herders from different ecogeographic zones during this early contact was integral to the formation, rise and expansion of pastoralist groups around 3300 BC. Main During the fifth and fourth millennia BC, key technological and social changes took place in southeastern Europe (SEE) which profoundly transformed prehistoric societies. Metal production was among the most important innovations; copper was mined, smelted and used to make axes, jewellery and small tools. The discovery of the necropolis of Varna (4600–4300 BC) on the Black Sea coast led to a reassessment of social inequality in human prehistory, with large quantities of gold and other symbols of power and wealth suggesting unprecedented levels of social stratification10,11,12. The many tell settlements that emerged during the Copper Age (CA, 4900–3800 BC) in SEE, involved in the proto-industrial exploitation of copper13, gold and salt, highlight this advanced social organization and the blossoming of social, political, economic and artisanal activities. Eminent tell sites include Mound Măgura Gorgana near Pietrele on the Lower Danube in Romania14, associated with the Gumelniţa culture and Tell Yunatsite in Bulgaria, associated with the Karanovo culture (Fig. 1 and Extended Data Fig. 1), which were occupied for several centuries15. From around 4600 BC, the similarity and continuous development of material culture and exchange of raw materials in the so-called Gumelniţa–Kodžadermen–Karanovo VI complex across southern Romania (Gumelniţa), northern Bulgaria (Kodžadermen) and Thrace (Karanovo) indicate transregional connectedness and suggest a relatively stable sociopolitical network. Consequently, the roughly simultaneous abandonment of the numerous tell settlements and cemeteries around 4250/4200 BC appears enigmatic (Fig. 1a,c). The underlying circumstances are unclear and might have involved the depletion of resources, the deterioration of soils and possibly also violent conflicts, as evidenced by the destruction horizon at Tell Yunatsite16,17. Historically, this demise was associated with the arrival of new groups from the steppe18 but this proposal has lacked sufficient evidence. However, settlement activity over the following centuries was scarce in the entire western Black Sea region, indicating a ‘dark’ millennium with, for example, Yunatsite not being resettled until about 1,000 years later during the Early Bronze Age (EBA)15. Fig. 1: Geographical locations, genetic analyses and chronology of newly reported ancient Copper Age, Eneolithic and Early Bronze Age individuals. 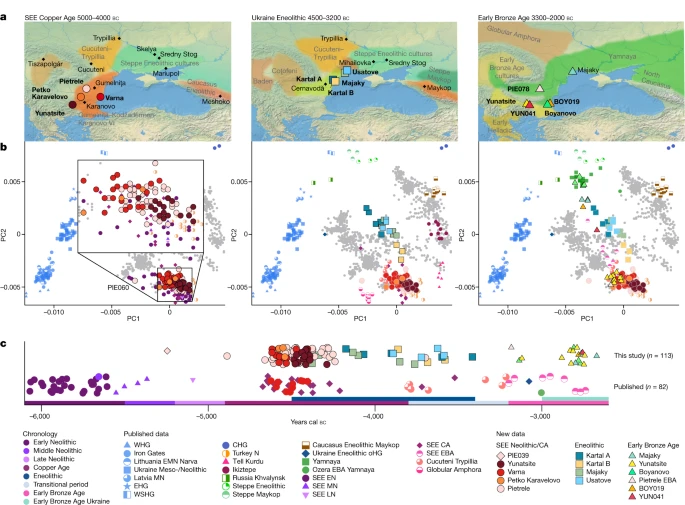 a, Map of sites and relevant archaeological cultures discussed in the text. Maps were made with Natural Earth (https://naturalearthdata.com). b, PCA of newly reported individuals (coloured symbols with black outline) and relevant published groups (coloured symbols, no outline) projected onto the West Eurasian genetic variation of 1,253 individuals from 77 populations. c, Mean radiocarbon dates of relevant published and newly reported individuals from southeastern Europe plotted according to the regional chronology. The suffixes in the group labels present archaeological time periods and geographical regions: N, Neolithic; EN, MN, LN, Early, Middle, Late Neolithic, respectively; CA, Copper Age; EBA, Early Bronze Age; SEE, southeastern Europe; WHG, EHG, WSHG, CHG, oHG, Western, Eastern, West Siberian, Caucasus, outlier Hunter–Gatherers, respectively. Following the end of the CA, the centre of settlement activity shifted further northeast towards the forest–steppe region, where huge settlements, with thousands of houses, the so-called megasites of the Cucuteni–Trypillia complex (around 4100–3800 BC), emerged19,20. This northwestern Black Sea region represents an interaction zone between late CA farming-associated groups and those of the adjacent steppe region with different ecogeographic conditions. Continued innovations increased human mobility and the exploration of lands hitherto not amenable to agrarian lifestyles, as practised in the regions in SEE and south of the Caucasus for many millennia before. From the contact zones in the northwestern Black Sea region and the Caucasus, a gradual transition from foraging to semi-nomadic pastoralism also followed in the North Pontic region during the sixth and fourth millennium BC, triggered by continued innovations6, transfer of livestock and advances in herd management, food processing, dairying practices8,9,19 and the development of arsenical-copper alloys21. The North Pontic region played a central role in the development of the oldest wheeled vehicles22, while the North Caucasian Maykop culture was critical in the further development of metal alloys, as well as early horse domestication8,9 and a sheep-wool economy combined with extensive dairy production9. The Maykop culture had extremely rich burials with metal weapons indicating ‘high status’ individuals, attesting to social inequality and upheaval during this time23, as such social elites were also found in southern Romania and Bulgaria24. The Cernavodă I (around 4000–3200 BC) and Usatove cultures (3600/3500–3200/3100 BC) in the northwestern Black Sea region played a chief role in the east–west exchange between the Danube and the lower Dniepr25 and these formations, while presumably indigenous, received strong contributions from the Trypillian tradition26. Similar to the SEE CA tell sites, the megasites and cultural phenomena of the northwestern Pontic region suddenly disappeared and were succeeded around 3300 BC by fully established pastoralists associated with the Yamnaya cultural complex. The expansion of North Pontic pastoralists to the west has been studied in many regions of Europe in recent years27, whereas their emergence and impact on societies in SEE is barely understood. This is relevant insofar as the archaeological record for the subsequent EBA (around 3200–2500 BC), indicates a concomitant rise in settlement activity for the first time since the demise of the CA settlements in the eastern Balkan region28. Burial mounds, associated with the Yamnaya cultural complex, appear frequently and extend along the Danube valley into the Carpathian Basin during the third millennium BC (refs. 4,29). By contrast, the resettlement of sites like Tell Yunatsite involved groups with burial rites not associated with the incoming steppe groups30 (Extended Data Fig. 1c,h). Archaeogenomic studies have shown that SEE CA individuals carry genetic profiles that resemble those of Neolithic farmers expanding from western Anatolia into Europe31, distinct from both the earlier pre-agropastoralist (Western and Eastern Hunter–Gatherers; WHG/EHG) and later EBA pastoralist groups1,2,4,5, who carried ‘steppe’ ancestry. Individuals from well-known, contemporaneous CA settlements (Pietrele and Yunatsite) and outstanding burial sites (Varna) provide a unique opportunity to study the genetic variation in and between sites at their peak settlement densities. However, the developments following early interactions, which had later given rise to the expansion of pastoralists and their genetic ancestry across Europe, remain unknown. Critically, individuals from the key period of the fifth and fourth millennium BC from the contact zone between SEE, the Trypillian megasites and the steppes have not been analysed genetically. Here, we address this spatial and temporal sampling gap by studying individuals associated with the Cernavodă I and Usatove cultures from the northwestern Black Sea region in today’s Ukraine. Additionally, we analyse EBA individuals from the tell sites Yunatsite and Pietrele, following a possible resettlement of the sites after several centuries of abandonment. We compare these to Yamnaya-associated individuals from eastern Bulgaria, who were buried in mounds typically associated with steppe pastoralists during the third millennium BC and to individuals postdating the Usatove horizon in the northwestern Black Sea area. In total, we report genome-wide data for 135 (out of 216 attempted) individuals from eight distinct sites (Fig. 1) ranging from around 5400 to 2400 BC: Neolithic (n = 1), CA (n = 95), Eneolithic (n = 18) and EBA (n = 21). All samples were enriched for a panel of 1.24 million single-nucleotide polymorphisms (1,240,000 SNP panel32), ranging from 61,000 to 947,000 SNPs with an average SNP coverage between 0.01× and 3.4×. We used a cut-off of 400,000 SNPs for hapROH and imputation and filtered for >550,000 SNPs for identity-by-descent (IBD) analyses (Supplementary Table A; Methods). We also report 113 new radiocarbon dates (Fig. 1c and Supplementary Table A). To assess the genetic ancestry and variation of the newly typed individuals we first performed principal component analysis (PCA) constructed from 1,253 modern-day West Eurasians from 77 different populations, onto which data from the ancient individuals were projected (Fig. 1b and Supplementary Table B; Methods). www.nature.com/articles/s41586-023-06334-8 |
|
|
|
Post by Admin on Jul 23, 2023 19:45:50 GMT
Neolithic and Copper Age ancestries The earliest-dated individual in our dataset, PIE039 from Pietrele, falls in the expected range of other SEE Neolithic individuals in PCA space, with whom she also shares affinities according to outgroup f3 statistics (Fig. 1b, Fig. 2 and Supplementary Table C). We used f4 statistics of the form f4 (test, PIE039; HGs, Mbuti), where ‘test’ are different Neolithic groups, to identify the genetically most similar Neolithic groups, which were then used as local proxies for quantitative ancestry modelling. We found Hungary_LN_Sopot and Malak Preslavets N to be most symmetrically related to PIE039 with respect to all HG comparisons (|Z| ≤ 1) and thus combined them into local group SEE 1, which could be used as a single source for proximal qpAdm modelling (P = 0.41), confirming shared local ancestry (Extended Data Fig. 2, Fig. 3d, Supplementary Tables D, E, H and Supplementary Information 5). Fig. 2: Outgroup f3 statistics for SEE CA, Ukraine Eneolithic and Early Bronze Age groups. 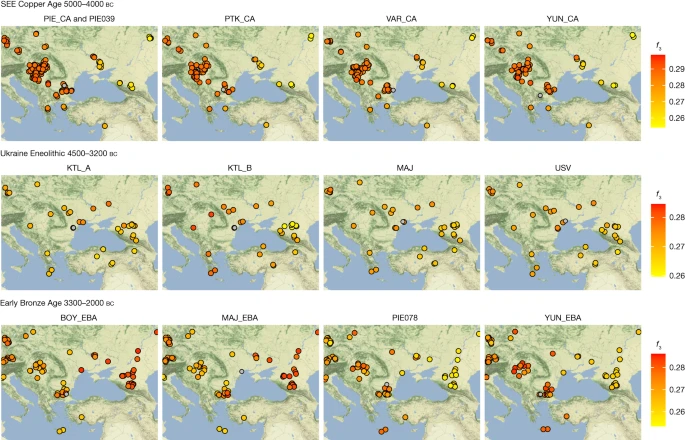 Outgroup f3 statistics of the form f3(test1, test2; Mbuti) plotted onto maps of central/SEE highlighting the shifting genetic affinities over time, separated temporally. Test1 includes groups and individuals newly reported in this study (headers) and their respective location is given by a grey circle. Test2 includes all relevant ancient populations from the respective time period (Supplementary Table C) and their locations are indicated as filled circles with black outlines. Higher f3 statistics (red colours) indicate more shared drift with the respective group in Test1. All f3 statistics, including outlier individuals and published ancient populations further west, can be found in Supplementary Table C. The maps were made in R57 and the tile sets are copyright of Stamen Design, under a Creative Commons Attribution (CC BY 3.0) licence. Fig. 3: Distal and proximal qpAdm results for the Copper Age, Ukraine Eneolithic and Early Bronze Age groups. 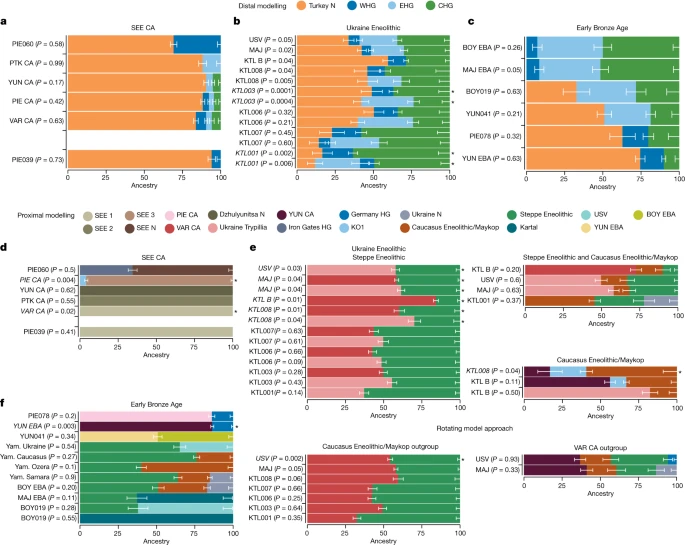 a–c, Distal models with Turkey_N, WHG, EHG and CHG as sources for the three sampled time periods: SEE CA (a); Ukraine Eneolithic (b); and Early Bronze Age (c). d–f, Geographically and temporally proximal models of the three sampled time periods: SEE CA (d); Ukraine Eneolithic (e); and Early Bronze Age (f) (Supplementary Tables G, H, P, Q, V and W). All results shown here were run with the parameter ‘allSNPs: NO’ (Supplementary Information 6). * Indicates non-supported/rejected/failed models when applying a P value cut-off of less than 0.05 (shown in italics). Ancestry proportions are shown with one standard error. Standard errors were computed with the default block jackknife approach. In PCA space, the chronologically younger SEE CA individuals from the emblematic sites of Yunatsite (YUN), Varna (VAR), Pietrele (PIE) and the multiple burial from Tell Petko Karavelovo (PTK), form a tight cluster that also overlaps with published Neolithic individuals from Anatolia and SEE29 (Fig. 1b). Moreover, outgroup f3 statistics suggest local genetic homogeneity throughout the CA in this region (Fig. 2 and Supplementary Table C). However, all SEE CA groups are slightly shifted towards the EHG/WHG cline in both PC1 and PC2 compared to most published Neolithic individuals. Distal qpAdm modelling (Fig. 3a and Supplementary Table G) confirmed minimal amounts of EHG-, CHG- and WHG-like ancestry, in addition to predominantly Turkey_N-like ancestry. This ancestry composition is already present during the Neolithic29 and confirmed by the test f4 (test, CA; HGs, Mbuti) in which Neolithic groups form a clade with SEE CA with respect to HG groups (Extended Data Fig. 2, Supplementary Tables D and E and Supplementary Information 5). This allows us to identify the best local Neolithic proxy for each SEE CA group and to account for the subtle differences in ancestries. Using the respective, locally preceding, Neolithic groups for proximal qpAdm modelling, we could model all SEE CA groups as a single-source model (Fig. 3d and Supplementary Table H), suggesting genetic continuity at the local scale. The outlier individual PIE060 is shifted further towards the WHG/EHG cluster in PCA, suggesting an excess of this type of ancestry, which could be confirmed by f4 statistics of the form f4(SEE N, PIE060; HGs, Mbuti) ((|Z| ≥ 3); Supplementary Table F). Ancestry modelling with qpAdm supports a two-way model (Fig. 3d) with SEE N (around 65%) and Iron Gates HG or KO1 (around 35%) as the best proxies. Using DATES33 to determine the time of admixture between SEE N and Iron Gates HG as a local HG ancestry, we obtained an admixture estimate of 16.3 ± 13.4 generations (Z = 1.213), which corresponds to around 81–832 years before the mean 14C date of PIE060, when a generation time of 28 years is assumed34. A flat decay curve (Extended Data Fig. 3a) supports the interpretation of a recent admixture date, which suggests that PIE060 came from a community outside Pietrele with recent contact with HGs. Indeed, individuals with similarly high amounts of HG ancestry have been reported from nearby sites in Malak Preslavets (around 70 km) and Dzhulyunitsa (around 140 km)29.
Poseidon_ID Culture Period Country Location Date (BP) mtDNA Y-DNA Terminal SNP
PIE039 Dudeşti Neolithic Romania Giurgiu County 6274 T2c1d1
PIE009 Gumelnița CA Romania Giurgiu County 5686 H A A
PIE005 Gumelnița CA Romania Giurgiu County 5699 K1a+195 C1a2b Z38888
PIE050 Gumelnița CA Romania Giurgiu County 5664 H11 C1a2b Z38888
PIE057 Gumelnița CA Romania Giurgiu County 5589 HV+16311 C1a2b Z38888
PIE063 Gumelnița CA Romania Giurgiu County 5657 I5a C1a2b Z38888
PIE076 Gumelnița CA Romania Giurgiu County 5437 H2 C1a2b Z38888
PIE032 Gumelnița CA Romania Giurgiu County 5574 T2b G2a P15/PF3112
PIE062 Gumelnița CA Romania Giurgiu County 5441 H G2a P15/PF3112
PIE022 Gumelnița CA Romania Giurgiu County 5531 W3b G2a2a PF3147
PIE033 Gumelnița CA Romania Giurgiu County 5734 X2i+@225 G2a2a1a2a FGC7739/Z6488
PIE054 Gumelnița CA Romania Giurgiu County 5424 I5 G2a2b2a P303/Page108/PF3340/S135/Z765
PIE058 Gumelnița CA Romania Giurgiu County 5421 I5 G2a2b2a1 P303/Page108/PF3340/S135/Z765
PIE015 Gumelnița CA Romania Giurgiu County 5704 V1a G2a2b2a1a1c2 FGC12126
PIE029 Gumelnița CA Romania Giurgiu County 5696 K1a+195 G2a2b2b PF3359
PIE037 Gumelnița CA Romania Giurgiu County 5439 N1a1a1a3 H2 P96
PIE060 Gumelnița CA Romania Giurgiu County 5813 H+152 I2a1b1a2a L701
PIE061 Gumelnița CA Romania Giurgiu County 5698 H+152 I2a1b1a2a L701
PIE007 Cernavoda? CA Romania Giurgiu County 4933 K1a2 J2a M410/PF4941
PIE028 Gumelnița CA Romania Giurgiu County 5600 K1a J2a1 PF4610
PIE044 Gumelnița CA Romania Giurgiu County 5614 - J2a1 PF4610
PIE026 Gumelnița CA Romania Giurgiu County 5651 U3a1 R1b1 L754/PF6269/YSC0000022
PIE064 Gumelnița CA Romania Giurgiu County 5672 K1a1b R1b1a1b M269/PF6517
PIE004 Gumelnița CA Romania Giurgiu County 5770 H R1b1b V88
PIE013 Gumelnița CA Romania Giurgiu County 5623 H R1b1b V88
PIE017 Gumelnița CA Romania Giurgiu County 5761 J1 R1b1b V88
PIE019 Gumelnița CA Romania Giurgiu County no collagen T2b R1b1b V88
PIE023 Gumelnița CA Romania Giurgiu County 5676 J2b1 R1b1b V88
PIE042 Gumelnița CA Romania Giurgiu County 5638 T2 R1b1b V88
PIE025 Gumelnița CA Romania Giurgiu County 5678 H5 T1 L206
PIE016 Cernavoda? CA Romania Giurgiu County K1a2
PIE031 Cernavoda? CA Romania Giurgiu County
PIE003 Gumelnița CA Romania Giurgiu County 5586 K1a24
PIE006 Gumelnița CA Romania Giurgiu County 5666 T2+16189
PIE008 Gumelnița CA Romania Giurgiu County 5593 T2b
PIE010 Gumelnița CA Romania Giurgiu County 5485 K1a
PIE012 Gumelnița CA Romania Giurgiu County 5705 H7
PIE014 Gumelnița CA Romania Giurgiu County 5641 H7
PIE018 Gumelnița CA Romania Giurgiu County 5602 -
PIE030 Gumelnița CA Romania Giurgiu County 5419 T2c1d1
PIE034 Gumelnița CA Romania Giurgiu County 5741 H5
PIE035 Gumelnița CA Romania Giurgiu County 5700 K1
PIE038 Gumelnița CA Romania Giurgiu County 5722 K1a
PIE041 Gumelnița CA Romania Giurgiu County 5706 K1a+195
PIE043 Gumelnița CA Romania Giurgiu County 5996 J1c15
PIE048 Gumelnița CA Romania Giurgiu County 5728 H4a1
PIE053 Gumelnița CA Romania Giurgiu County 5599 H11b
PIE064 Gumelnița CA Romania Giurgiu County 5672 K1a1b
PIE065 Gumelnița CA Romania Giurgiu County 5721 K1a4
PIE069 Gumelnița CA Romania Giurgiu County 5683 H
PIE074 Gumelnița CA Romania Giurgiu County 5475 T2c1d+152
PIE075 Gumelnița CA Romania Giurgiu County 5508 H
PIE077 Gumelnița CA Romania Giurgiu County 5411 T2b+152
MAJ003 Usatove CA Ukraine Odessa Oblast 4978 U5a1a1 G2a2b2a G-P303
UBK006 Usatove CA Ukraine Odessa Oblast 4853 U4a2 I2a1a I-P37.2
KTL006 Cernavodă I CA Ukraine Odessa Oblast 5205 U5a1a I2a1b1a2a2a I-L699
KTL001 Cernavodă I CA Ukraine Odessa Oblast 4609 H2a1 I2a1b1a2a2a2 I-S12195
KTL005 Cernavodă I CA Ukraine Odessa Oblast 4905 U3b3 R1b R-M343
MAJ009 Usatove CA Ukraine Odessa Oblast 5545 H5 R1b1a2a2 R-L754
KTL002 Cernavodă I CA Ukraine Odessa Oblast 4806 J1c5
KTL003 Cernavodă I CA Ukraine Odessa Oblast 4840 T2c1c1
KTL004 Cernavodă I CA Ukraine Odessa Oblast 5064 T2b
KTL007 Cernavodă I CA Ukraine Odessa Oblast 5104 U4
KTL008 Cernavodă I CA Ukraine Odessa Oblast 4895 U2e1b2
MAJ002 Usatove CA Ukraine Odessa Oblast 5498 I1b*1
MAJ008 Usatove CA Ukraine Odessa Oblast 5270 I1a1
MAJ020 Usatove CA Ukraine Odessa Oblast 5058 U5a1f1
MAJ023 Usatove CA Ukraine Odessa Oblast 5311 I5
UBK003 Usatove CA Ukraine Odessa Oblast 5110 U5a1f1
USV004 Usatove CA Ukraine Odessa Oblast 4914
USV005 Usatove CA Ukraine Odessa Oblast 4819 X2b
YUN013 Gumelnița/Karanovo CA Bulgaria Pazardzhik 5624 H7 C1a2b Z38888
YUN015 Gumelnița/Karanovo CA Bulgaria Pazardzhik 5541 HV+16311 G2a2b L30/PF3267/S126
YUN009 Gumelnița/Karanovo CA Bulgaria Pazardzhik 5548 H62 G2a2b2b1a2 FGC82036
YUN048 Gumelnita/Karanovo CA Bulgaria Pazardzhik 5657 K1a1 H2 P96
YUN026 Gumelnița/Karanovo CA Bulgaria Pazardzhik 5637 T2e J2a1a2b2~ FGC8506
YUN007 Gumelnița/Karanovo CA Bulgaria Pazardzhik 5551 H J2a1a2b2a2~ SK1359/Y7010/Z27851
YUN049 Gumelnita/Karanovo CA Bulgaria Pazardzhik
YUN001 Gumelnița/Karanovo CA Bulgaria Pazardzhik 5614 K2a
YUN003 Gumelnița/Karanovo CA Bulgaria Pazardzhik U2+152
YUN005 Gumelnița/Karanovo CA Bulgaria Pazardzhik 5570 U4a2
YUN006 Gumelnița/Karanovo CA Bulgaria Pazardzhik 5589 K1a
YUN012 Gumelnița/Karanovo CA Bulgaria Pazardzhik X2b
YUN014 Gumelnița/Karanovo CA Bulgaria Pazardzhik 5577 T2b23
YUN019 Gumelnița/Karanovo CA Bulgaria Pazardzhik U8b1a2b
YUN021 Gumelnița/Karanovo CA Bulgaria Pazardzhik 5431 K1a2
YUN025 Gumelnița/Karanovo CA Bulgaria Pazardzhik X2b6
YUN027_I0781.merge Gumelnița/Karanovo CA Bulgaria Pazardzhik K1a+195
YUN028 Gumelnița/Karanovo CA Bulgaria Pazardzhik 5625 H
YUN031 Gumelnița/Karanovo CA Bulgaria Pazardzhik 5652 K1a
PTK005 Chalcolithic, Karanovo VI, Gumelnita CA Bulgaria Polski Tyrambesh Municipality U2+152 G2a2b2a1a1c1a1 CTS5990/Z1903
PTK001 Chalcolithic, Karanovo VI, Gumelnita CA Bulgaria Polski Tyrambesh Municipality 5709 HV+16311 I2a1b M436/P214/PF3856/S33
PTK004 Chalcolithic, Karanovo VI, Gumelnita CA Bulgaria Polski Tyrambesh Municipality 5708 H3aq I2a1b M436/P214/PF3856/S33
PTK003.merged Chalcolithic, Karanovo VI, Gumelnita CA Bulgaria Polski Tyrambesh Municipality 5721 H3aq R1b1b V88
VAR009 Varna CA Bulgaria Varna 5659 K1a1 E1b1b1a1b1 L618
VAR018 Varna CA Bulgaria Varna 5608 T2c1d1 E1b1b1a1b1 L618
VAR004 Varna CA Bulgaria Varna 5732 J2b1 G2a2a1 PF3148
VAR011 Varna CA Bulgaria Varna 5574 H G2a2a1 PF3148
VAR023_ANI160 Varna CA Bulgaria Varna 5735 H G2a2b2b PF3359
VAR030 Varna CA Bulgaria Varna 5650 K2a I2a1b1a2a~ L701
VAR006 Varna CA Bulgaria Varna 5600 J1c I2a2 L596/PF6907/S292
VAR008 Varna CA Bulgaria Varna 5765 T2b R1b1b V88
VAR019 Varna CA Bulgaria Varna 5773 H R1b1b V88
VAR020 Varna CA Bulgaria Varna 5687 U5b1b1a R1b1b V88
VAR026 Varna CA Bulgaria Varna 5668 H R1b1b V88
VAR016 Varna CA Bulgaria Varna 5631 T1a T1a M70
VAR010 Varna CA Bulgaria Varna 5749 U5b1 T1a2 L131
VAR022 Varna CA Bulgaria Varna 5677 H1e T1a2b L446
VAR032 Varna CA Bulgaria Varna 5661 K1a19 T1a2b L446
VAR002 Varna CA Bulgaria Varna 5672 X2
VAR003 Varna CA Bulgaria Varna H
VAR012 Varna CA Bulgaria Varna 5689 T2b28
VAR017 Varna CA Bulgaria Varna 5714 K1c
VAR021 Varna CA Bulgaria Varna 5661 K1a2
VAR027 Varna CA Bulgaria Varna 5721 K1
VAR033 Varna CA Bulgaria Varna 5787 U1a1d
VAR034 Varna CA Bulgaria Varna 5740 H87
PIE078 Gumelnița EBA Romania Giurgiu County 4463 K1a1a I2a1a2b L621/S392
MAJ017 Usatove EBA Ukraine Odessa Oblast 4242 H2a3 R1b1a1b1b R-Z2103
MAJ004 Usatove EBA Ukraine Odessa Oblast 4107 U5b2a1a1
MAJ019 Usatove EBA Ukraine Odessa Oblast 4185 H1
YUN038 EBA EBA Bulgaria Pazardzhik 4248 W1h I2a1a2b1 CTS10936
YUN037 EBA EBA Bulgaria Pazardzhik W1h I2a1a2b1a CTS4002
YUN033 EBA EBA Bulgaria Pazardzhik 4428 T2e2a
YUN034 EBA EBA Bulgaria Pazardzhik 4239 N1a1a1a
YUN035 EBA EBA Bulgaria Pazardzhik T2b
YUN036 EBA EBA Bulgaria Pazardzhik 4167 T2b
YUN039 EBA EBA Bulgaria Pazardzhik 4213 H11a
YUN041 EBA EBA Bulgaria Pazardzhik 4161 K1c1
YUN042 EBA EBA Bulgaria Pazardzhik 4181 H
YUN043 EBA EBA Bulgaria Pazardzhik 4162 K1a3a
YUN044 EBA EBA Bulgaria Pazardzhik no collagen H1
YUN045 EBA EBA Bulgaria Pazardzhik 4270 T2b
BOY001 Yamnaya EBA Bulgaria Yambol Oblast 4205 R1a1a R1b1a1 P297/PF6398
BOY008 EBA (extended) EBA Bulgaria Yambol Oblast 4213 H R1b1a1b M269
BOY009 EBA (crouched) EBA Bulgaria Yambol Oblast 4218 H13a1a1 R1b1a1b1b3a Z2109
BOY019 EBA (extended) EBA Bulgaria Yambol Oblast 4212 U5a2b R1b1a1b1b3a Z2109
BOY014 EBA (extended) EBA Bulgaria Yambol Oblast 4421 U5a1b
In line with the autosomal data, the Y-chromosomal and mitochondrial DNA lineages are common in nearly all Neolithic and CA groups studied until now, albeit with several males also carrying typical Mesolithic (C1a and I2a) Y lineages35, including individual PIE060 (Extended Data Fig. 3b and Supplementary Table A). With seven different main lineages among 29 males in Pietrele (I2a1, C1a, G2a, H2, T1a, J2a and R1b-V88), six among 15 males in Varna (I2a1, I2a2, G2a, T1a, E1b1 and R1b-V88) and four among six males at Yunatsite (C1a, G2a, H2, J2a), the Y-chromosomal diversity during the SEE CA was higher than in central/western Europe36,37,38. When testing for genetic relatedness in each of the SEE CA sites using READ, we detected only three first-degree and two second-degree relationships in total (Supplementary Table I; Methods). To specifically test for links between the contemporaneous SEE CA sites and for more distant genetic relatedness we explored signals of IBD sharing between individuals in and between all sites (Methods). We found no evidence for between-site links up to the fourth to fifth degree and only two pairs of individuals (PIE003-VAR010 and YUN005-VAR030) shared at least two blocks greater than 20 cM indicative of a fifth to seventh degree relationship (Extended Data Fig. 4a and Supplementary Table J). Integrating the normalized sum and number of shared blocks we find higher background relatedness at the intrasite level at Yunatsite and Varna compared to Pietrele, which can be explained by the structure of the sites (a destruction horizon of households and a burial ground with shorter use, respectively, versus tell and settlement burials spanning 350–400 years) (Fig. 1c and Extended Data Fig. 4b). However, analysis of the runs of homozygosity (ROH) per individual using hapROH indicates low levels of parental background relatedness suggesting relatively large effective population sizes, consistent with previous observation across early farming societies (Methods; Extended Data Fig. 5). These findings reflect the settlement density and the wide-spread cultural, rather than close genetic, connectedness of the Gumelniţa–Kodžadermen–Karanovo VI complex, in line with the cross-regional significance of SEE tell sites26. |
|
|
|
Post by Admin on Jul 24, 2023 20:25:58 GMT
Early contacts during the Eneolithic
Eneolithic individuals from Ukraine (Ukraine Eneolithic), dated from around 4500–3500 BC, associated with the Cernavodă I and Usatove cultures, form a genetic cline in PCA space (Fig. 1b) between Neolithic/SEE CA individuals and published Eneolithic steppe individuals from the North Caucasus39 and Khvalynsk in western Russia32. This indicates possible admixture between CA farmer-related groups and Eneolithic steppe groups, as in line with cultural interactions described in the archaeological record40,41,42. The observed genetic cline reflects developments over a wide chronological range of around 1,000 years (Fig. 1c and Supplementary Table A). Some of the newly reported 14C dates could be affected by a freshwater reservoir effect43, common in Steppe Eneolithic sites44,45 and could therefore be several centuries younger than their reported dates. However, accounting for this possibility, an offset of around 500 years would still date most of the Ukraine Eneolithic individuals to the fourth millennium BC and thus considerably earlier than the Yamnaya-associated steppe pastoralist expansion.
Individuals from Kartal (around 4150–3400 BC), associated with the Cernavodă I culture, are genetically highly heterogeneous, with five individuals (Kartal A) forming a cline between ‘Steppe Eneolithic’/‘Steppe Maykop’ individuals and Early Neolithic groups, while three other individuals (Kartal B) fall closer to the latter (Supplementary Tables L and M). The five contemporaneous individuals from Majaky (MAJ), are genetically more homogeneous and fall together with the four individuals from the late Eneolithic Usatove type-site (USV/UBK; Supplementary Table A) in the middle of the ‘Kartal cline’. We tested for a correlation between positions of the Ukraine Eneolithic individuals in PC2 and their 14C dates and found none (Spearman’s ρ = 0.113, P = 0.6656). The broadscale shift in genetic affinities between the CA and the Eneolithic, from SEE to the steppe zone, is also clearly visible in outgroup f3 statistics when mapped geographically (Fig. 2 and Supplementary Table C).
To formally characterize the Ukraine Eneolithic individuals, we tested for excess shared ancestry with four Holocene ‘cornerstone’ populations (Turkey_N, WHG, EHG/WSHG and CHG) (Supplementary Information 1.2), using f4-symmetry statistics of the form f4(test, Ukraine Eneolithic; cornerstone, Mbuti) and conditioning on three test populations (Extended Data Fig. 6, Supplementary Tables E, M and N and Supplementary Information 6). First, compared to Turkey_N, Ukraine Eneolithic individuals show excess affinity to all HG groups, as indicated by significantly negative f4 statistics (|Z| ≥ 3) (Extended Data Fig. 6a). Second, conditioning on Steppe Eneolithic (Extended Data Fig. 6b), we observe excess affinity of Ukraine Eneolithic to Turkey_N, a symmetrical relatedness to CHG and WHG, while Steppe Eneolithic groups carry more EHG/WSHG ancestry. On the basis of cultural influences which also link the northern Black Sea through the steppe belt to the North Caucasus region40,41,42, we also test for potential influence of North Caucasian groups. Using Caucasus Eneolithic/Maykop as test (Extended Data Fig. 6c) we find excess affinity of Ukraine Eneolithic to EHG and WHG and Turkey_N, while Caucasus Eneolithic/Maykop share more drift with CHG.
The archaeological record identifies the northwestern Black Sea region as an interaction zone between late CA farming and groups from the steppe region19,20. Such an early interaction has been postulated by Immel et al.46, who have reported Yamnaya-related ancestry in individuals associated with the Cucuteni–Trypillia complex from today’s Moldova. However, on re-analysis of these data we find that this signal can be explained solely by an increase in especially EHG-rich ancestry (Supplementary Information 4 and Supplementary Table O).
To characterize the role of Cernavodă I and Usatove-associated individuals from the postulated interaction zone, who show a clear signal of admixture, we formally tested the contribution of diverse ancestry sources using f4(Steppe Eneolithic/Caucasus Eneolithic/Maykop, Ukraine Eneolithic; test, Mbuti), where test represents SEE and Anatolian CA farmer groups (Supplementary Table P). With respect to Steppe Eneolithic, all Ukraine Eneolithic individuals show excess affinity to all tested CA groups. With respect to Caucasus Eneolithic/Maykop, USV, MAJ, KTL_B, KTL003 and KTL008 show excess affinity to all SEE CA farmer groups, while KTL006 and KTL007 only share drift with Ukraine Trypillia (Supplementary Table P).
Of note, all f4-symmetry tests with Caucasus Eneolithic/Maykop and SEE CA indicate an additional attraction of Ukraine Eneolithic to WHG/EHG (Supplementary Table N), with Iron Gates HG or Ukraine N showing the highest affinity (Supplementary Table Q). This affinity towards WHG/EHG is absent when Steppe Eneolithic is used (Supplementary Table N), implying that scenarios involving potential gene flow from the Caucasus would require an additional source carrying WHG-/EHG-like ancestry as this ancestry is not sufficiently represented by SEE CA or Caucasus Maykop groups.
Using distal qpAdm modelling we find support for a four-way admixture of Turkey_N, EHG, CHG and WHG for KTL001, KTL007, MAJ and USV (Fig. 3b and Supplementary Table R), while individuals KTL003, KTL006 and KTL008 can be modelled alternatively with three sources (Turkey_N+EHG+CHG) and KTL_B individuals only with Turkey_N (around 60%), CHG (around 28%) and WHG (around 12%) ancestry. Following up with proximal qpAdm models to explore potential contribution(s) of temporally and geographically closer groups (Fig. 3e and Supplementary Tables E and S), we find that all Ukraine Eneolithic individuals can be modelled as a two-way model of either VAR_CA or Ukraine Trypillia as farmer-related ancestry source and Steppe Eneolithic as a source of mixed EHG+CHG ancestry.
Since archaeological research suggests a cultural contribution of Steppe Eneolithic and Maykop groups (Supplementary Information 2.2), we specifically tested for alternative scenarios which involved admixture between both groups north of the Caucasus and subsequent spread westwards. Using both associated ancestries and different HGs and SEE CA-related groups as sources in qpAdm modelling (Fig. 3e and Supplementary Table S), we find that KTL001 can indeed be modelled as a three-way mixture of Steppe Eneolithic (around 32%), Caucasus Eneolithic/Maykop (around 46%) and Ukraine N foragers (around 22%), to the exclusion of a SEE CA source. By contrast, MAJ and USV can be modelled as VAR_CA or Ukraine Trypillia (around 50%), Steppe Eneolithic (around 35%) and Caucasus Eneolithic/Maykop (around 15%) as minor third component. KTL_B results in the same model but with a higher VAR_CA component (around 73%) and a minor contribution of Steppe Eneolithic (around 10%) ancestry (Supplementary Table S).
Exploring an alternative scenario which excludes Steppe Eneolithic as a source, we find a well-fit model for KTL008 with YUN_CA (around 17%), Caucasus Eneolithic/Maykop (around 60%) and KO1 (around 23%). Further, KTL_B can be modelled with Ukraine Trypillia (around 82%) and Caucasus Eneolithic/Maykop (around 18%) as second source, which is consistent with the omission of EHG ancestry in the distal qpAdm results for KTL_B (Fig. 3e and Supplementary Table S).
Finally, to test whether we can distinguish between the farmer-related ancestry contributed by SEE CA- or Maykop-associated groups from the Caucasus, or by both, we rotated each source to the outgroups, alternatingly, keeping Steppe Eneolithic as a constant. Here, we find strong support for a genetic contribution from SEE CA rather than Caucasus Eneolithic/Maykop for most KTL individuals (except KTL_B), which can be modelled as Steppe Eneolithic and VAR_CA (Fig. 3e and Supplementary Table S). The same model is supported for MAJ (P = 0.05) but rejected for USV, which indicates that Maykop-associated ancestry is needed for the latter. Indeed, the competing model, with Maykop as an additional source and VAR_CA as an outgroup, results in a well-fit four-way mixture model for USV (P = 0.93) and improved model fit for MAJ (P = 0.33), whereas the models for the remaining KTL individuals are rejected (Supplementary Table S). This provides strong support for an alternative admixture history for USV and MAJ, involving local SEE CA, Steppe Eneolithic, Caucasus Eneolithic/Maykop and a HG-related source, a combination that is distinct from KTL individuals.
The similarities in genetic ancestry presented for MAJ and USV are also observed in the results from the IBD analysis (Extended Data Fig. 4a and Supplementary Table J) in which we find a fourth to sixth degree relationship between MAJ023 and USV006, which reflects the close geographical vicinity of the two sites. The normalized sum and number of shared blocks for Ukraine Eneolithic show a higher background relatedness in USV compared to the other sites (Extended Data Fig. 4b) but also between USV and MAJ and USV and KTL, respectively, which matches the relative chronological overlap of the three sites (Fig. 1c and Supplementary Table A). However, in comparison to the preceding CA and heterogenous KTL individuals, ROH indicate a slightly elevated parental background relatedness for MAJ and USV (Extended Data Fig. 5), suggesting smaller effective population sizes in Usatove-associated groups.
Y-chromosomal evidence from the six Ukraine Eneolithic males reflects lineages from each of the contributing sources (Extended Data Fig. 3b): G2a is probably a Neolithic legacy, while three males carrying I2a1 could be attributed to the local Ukrainian Neolithic or HG groups in general. KTL005 and MAJ009 carry haplotypes R1b/M343(×P297) and R1b1/L754(×M269), respectively, which are ancestral for the pre-M269 branch (P297) and the M269 branch. Importantly, we do not observe R1b-Z2103 or immediate R1b-M269 precursor lineages, which originated in the steppe and are later linked with expansion of steppe-related ancestry.
|
|
|
|
Post by Admin on Jul 25, 2023 19:41:39 GMT
Genetic ancestries during the Bronze Age
The EBA individuals in this study are characterized by two contrasting clusters of genetic ancestry in PCA space (Fig. 1b) and different genetic affinities in outgroup f3 statistics (Fig. 2 and Supplementary Table C). Individuals from YUN and individual PIE078, who date to the first half of the third millennium BC, resemble the SEE CA groups, whereas BOY_EBA and MAJ_EBA individuals fall within the ‘steppe ancestry’ cluster, commonly associated with the Yamnaya cultural complex. Two outlier individuals, BOY019 and YUN041, fall in the space between. Intriguingly, the males from YUN_EBA/PIE078 carried Y-chromosome lineages I2a, suggestive of a HG legacy, while the males from BOY/MAJ_EBA carried R1b-Z2103 or derived lineages, a characteristic hallmark of Yamnaya-associated ancestry (Extended Data Fig. 3b).
On the basis of these observations we tested for additional attraction towards HG-related groups in YUN_EBA and PIE078 compared to their CA predecessors by using f4(CA, EBA; HGs, Mbuti) and confirmed the excess HG ancestry in EBA individuals from YUN and PIE with significant negative results (|Z| ≤ 3) (Extended Data Fig. 7 and Supplementary Table T). By contrast, for MAJ_EBA, BOY_EBA, BOY019 and YUN041, we tested for additional attraction towards farmer-related groups represented by VAR_CA when compared to Yamnaya-associated groups (test) using f4(test, EBA, VAR_CA, Mbuti) (Supplementary Table E). Here, only the outlier individual YUN041 has a higher affinity to VAR_CA than to other EBA groups (Extended Data Fig. 8 and Supplementary Table U). Distal qpAdm modelling with cornerstone populations confirms the contrasting ancestries of the two main EBA clusters. PIE078 and YUN_EBA can be modelled with Turkey_N, CHG and WHG (Fig. 3c and Supplementary Table X), whereas MAJ_EBA, BOY_EBA, BOY019 and YUN041 require EHG ancestry as an additional source (Fig. 3c).
We then explored the apparent homogeneity of Yamnaya-associated EBA steppe pastoralist groups, by testing for possible contribution(s) from four sources: Ukraine Eneolithic as a proxy for mixed Turkey_N/CHG/EHG ancestry, Ukraine N as an HG-related group, Steppe Eneolithic as pre-Yamnaya genetic substrate and Caucasus Eneolithic/Maykop as a proxy for mixed Turkey_N/CHG-related South Caucasus ancestry, as suggested by ref. 47 and directly supported by our results for the preceding Eneolithic period. First, we formally tested for shared drift between all EBA Yamnaya-associated individuals and Steppe Eneolithic/Caucasus Eneolithic/Maykop with respect to cornerstone populations by using f4(Steppe Eneolithic/Caucasus Eneolithic/Maykop, EBA; cornerstones, Mbuti). With the exception of Yamnaya Caucasus, all EBA individuals show an excess affinity to Turkey_N when compared to Steppe Eneolithic (Extended Data Fig. 9 and Supplementary Table V). Further, when compared to Caucasus Eneolithic/Maykop all EBA individuals share drift with WHG and EHG/WSHG and only YUN041 is also significant for Turkey_N (Extended Data Fig. 9 and Supplementary Table V). Second, we used f4-symmetry statistics of the form f4(steppe1, steppe2; test, Mbuti) where test includes Ukraine N, Ukraine Eneolithic, Caucasus Eneolithic/Maykop and Steppe Eneolithic. Here, with the exception of outlier individual Ukraine_Ozera_EBA_Yamnaya, all f4 statistics are non-significant (|Z| ≤ 3) (Supplementary Table W), which indicates that all Yamnaya-associated individuals including those from Ukraine and Bulgaria are genetically highly similar.
Applying the same rationale and sources to proximal qpAdm modelling to uncover subtle signals (Fig. 3f and Supplementary Table Y), we find that BOY_EBA and Yamnaya Samara can be modelled as a three-way mixture of Steppe Eneolithic, Caucasus Eneolithic/Maykop and Ukraine N. We note that the same three sources contributed to the preceding Ukraine Eneolithic individuals from USV and MAJ (in addition to SEE CA ancestry), which suggests that similar processes had led to the tripartite ancestry formation in the steppe zone during the fourth millennium BC. Indeed, we find that BOY_EBA, MAY_EBA and Yamnaya Samara can also be modelled as a two-way mixture of Steppe Eneolithic and KTL001 (who lacked SEE_CA ancestry). For Ukraine_EBA_Yamnaya, we find support for a three-way model (P = 0.07) with Steppe Eneolithic (around 75%), Caucasus Eneolithic/Maykop (around 14%) and Globular Amphora (around 11%) as a western source but also improved model fit (P = 0.5) for a two-way mixture of Steppe Eneolithic (around 65%) and USV (around 35%) (Supplementary Table Y), which suggests a possible direct contribution of Ukraine Eneolithic groups to steppe pastoralists in the third millennium BC. By contrast, Yamnaya Caucasus individuals from the southern steppe can be modelled as a two-way model of around 76% Steppe Eneolithic and 26% Caucasus Eneolithic/Maykop, confirming the findings of Lazaridis and colleagues47. This two-way mix (40% + 60%, respectively) also provides a well-fit model (P = 0.09) for the Ozera outlier individual, consistent with the position in PCA and corroborating an influence from the Caucasus. Despite the overlap in PCA, these results suggest subtle geographical structure, involving local genetic strata and influences from neighbouring groups in western and southern contact zones, respectively. Individual BOY019 can be modelled successfully with around 63% USV and around 37% Steppe Eneolithic ancestry or around 40% Ukraine Trypillia and around 60% Steppe Eneolithic, suggesting interaction between these two neighbouring groups in the western contact zone or alternatively direct descent from admixed groups (for example, KTL001). Finally, individual YUN041 can be modelled as around 50% local YUN_EBA ancestry and 50% of either BOY_EBA or another Yamnaya-associated source.
|
|
|
|
Post by Admin on Jul 26, 2023 18:55:22 GMT
Discussion
The genetic homogeneity observed in and across the four CA sites (PIE, YUN, PTK and VAR) of the fifth millennium BC matches the cultural homogeneity of the archaeological records and suggests an extended period of a relative stable sociopolitical network and absence of large-scale cultural and genetic transformations. Shared shorter IBD tracts between sites are consistent with the transregional connectivity visible in the material culture. We can only speculate about the reasons that led to decreasing settlement densities at the end of the CA. Conflict arising from an early expansion of supposedly ‘Indo-European’ groups from the steppe, an idea that was put forward by M. Gimbutas18, is possible but internal competition and strife between CA groups is equally likely. In fact, given the near-identical genetic ancestry profiles of SEE CA groups, we caution that genetic analyses would be blind to internal conflicts, causing the replacement of one CA group by another. Long-lasting droughts and forest fires16 or infectious diseases and ensuing epidemics are other factors that could deplete lands. Indeed, evidence for early forms of Yersinia pestis as old as 5,000 years has been reported48,49,50 and even further back in time for Salmonella enterica51 for individuals associated with transitional foraging and pastoralism. Despite the systematic screening of teeth, we found no evidence for pathogens among the CA individuals of the fifth and fourth millennium BC, apart from two individuals (YUN048 and VAR021), who were positive for the Hepatitis B virus (HBV)52, while individual VAR021 was also positive for Salmonella enterica.
A principal finding from our study indicates early contact and admixture between CA farming groups from SEE and Eneolithic groups from the steppe zone in today’s southern Ukraine, possibly starting in the middle of the fifth millennium BC when settlement densities shifted further north, connecting the lower Danube region with the coastal steppe and Cucuteni–Trypillia groups of the forest–steppe. Archaeological evidence shows that the early CA Gumelniţa groups had already settled deep into the steppe zone by the mid-fifth millennium BC, introducing elements of a farming lifestyle but also carrying cultural influences from local HG groups53. The succeeding Cernavodă I and Usatove archaeological cultures were heavily influenced by local CA cultures and surrounds. During the fourth millennium BC, the northwestern Pontic region experienced intensified contact with Steppe Eneolithic groups, while these in turn also had contact with groups in the North Caucasus, such as Maykop, all of which are mirrored by the genomic data presented here. Moreover, despite the close geographical proximity of the Ukrainian sites studied, we were able to trace different admixture histories. Here, the heterogeneity of the individuals from the site Kartal stands out, which is located on the Danube delta at the northern end of the former distribution of the Chalcolithic Gumelniţa–Kodžadermen–Karanovo VI complex and thus represents the transformative nature and dynamics of the fourth millennium BC in action. By contrast, the more homogenous Majaky and Usatove groups, located north of the Dniester River, show that such assimilation processes had already occurred, suggesting that contact and exchange between transitional foragers and early pastoralist groups from the forest–steppe zone and non-local SEE farmer-associated groups had started already in the late fifth millennium BC. Moreover, variable cultural influences attested by the archaeological record40,41,53 are also traceable genetically. We argue that livestock, innovations and technological advances were exchanged through these zones of interaction, which then led to the establishment of fully developed pastoralism in the steppe by the end of the fourth millennium BC. Gene flow from both contact zones into the steppe could also explain the small amounts of farmer-related ancestry in the emerging Yamnaya pastoralists, which differentiates them from the Steppe Eneolithic substrate and accounts for subtle geographical structure in the vastly expanding territory/range.
The early admixture during the Eneolithic presented in this study appears to be local to the northwestern Black Sea region of the fourth millennium BC and did not affect the hinterland in SEE. In fact, EBA individuals from the fourth and third millennia BC from YUN and PIE do not show traces of steppe-like ancestry but instead a resurgence of HG ancestry observed widely in Europe during the fourth millennium BC (refs. 4,29,54,55). This indicates the presence of remnant HG groups in various non-farmed regions, for example, highlands and uplands or densely forested zones and wetlands and a mosaic of ancestries rather than a genetically uniform CA and EBA Europe.
While only a few tell sites have been resettled by local and/or incoming groups who did not originate in the North Pontic region, we can trace the appearance of migrants from the steppe, clearly attributed to Yamnaya culturally and genetically, in the local time transect at Majaky but also at Boyanovo in the Bulgarian lowlands of the Thracian Plain. The subtle differences in genetic ancestries between these two when compared to different Yamnaya-associated groups account for their geographical locations and different stages of genetic and perhaps, cultural assimilation. Two outlier individuals from EBA YUN and BOY bear witness to occasional admixture between inhabitants of EBA tells and incoming steppe pastoralists. Ultimately, the third millennium BC form of ‘steppe’-ancestry is expected to have reached the Great Hungarian plain, from where it diversified and spread further west. The interaction between local and incoming groups in SEE did not result in archaeologically visible conflicts or a near-complete autosomal genetic turnover as observed in Britain or a replacement of the Y-chromosome lineages in the Iberian Peninsula36,56.
Further integrated archaeogenomic studies are needed to disentangle the dynamics at play around the Black Sea during the formative periods of the admixture clines demonstrated in this study. High-quality genome-wide data from the fifth and fourth millennia BC that allow the direct tracing of IBD blocks shared by contributing groups will hold the key to understanding the population history of West Eurasia.
|
|

