|
|
Post by Admin on Jul 1, 2018 18:46:15 GMT
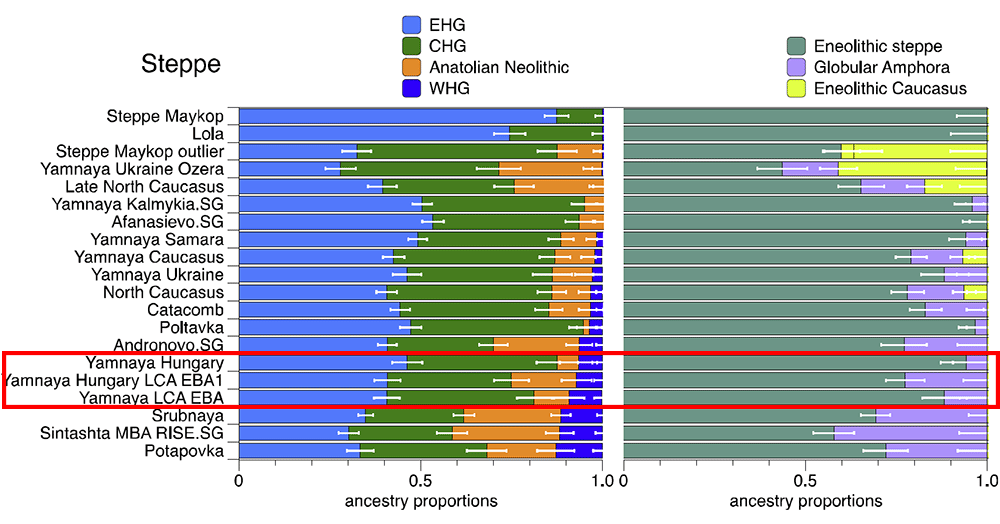
 Our results show that at the time of the eponymous grave mound of Maykop, the North Caucasus piedmont region was genetically connected to the south. Even without direct ancient DNA data from northern Mesopotamia, the new genetic evidence suggests an increased assimilation of Chalcolithic individuals from Iran, Anatolia and Armenia and those of the Eneolithic Caucasus during 6000-4000 calBCE23, and thus likely also intensified cultural connections. Within this sphere of interaction, it is possible that cultural influences and continuous subtle gene flow from the south formed the basis of Maykop. Also, unlike more recent times, the North Caucasian piedmont and foothill of the Caucasus region was more strongly connected to Northern Iran than to the steppe, at least until the Bronze Age. 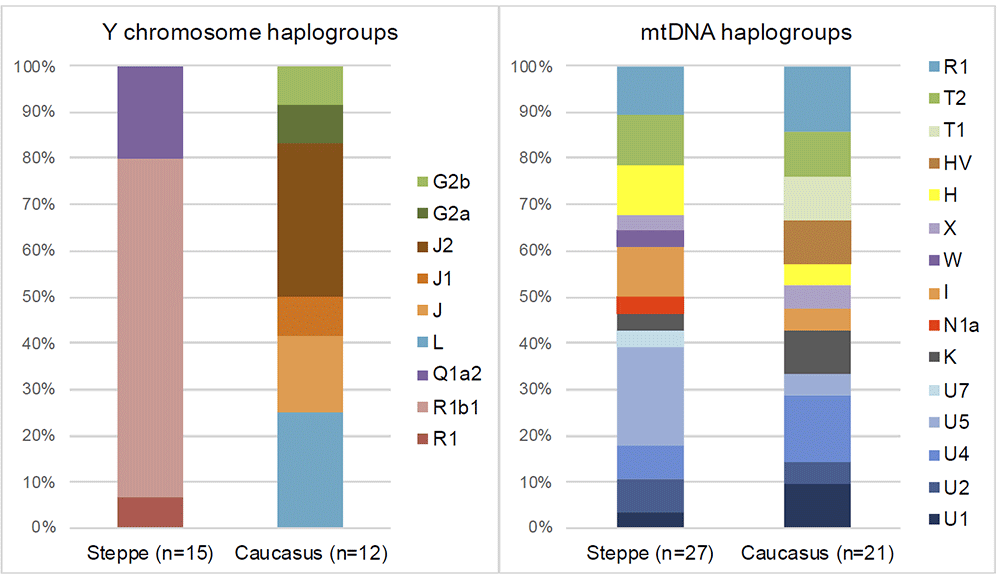 (…) our data shows that the northern flanks were consistently linked to the Near East and had received multiple streams of gene flow from the south, as seen e.g. during the Maykop, Kura-Araxes and late phase of the North Caucasus culture. Alternatively, R1b1 was plausibly native to the Pontic-Caspian steppes. A male from the Mesolithic Samara culture (5,650–5,555 BCE) belonged to R1b1, which was the earliest documented ancient sample of this haplogroup discovered to date (Haak et al. 2014). There was cultural interaction between the steppe herders and the Maykop people from the Near East (Wang et al. 2018) and the steppe herders could have adapted to the Maykop culture, thus abandoning their previous hunter-gatherer lifestyle. In this complex civilizing process, dynastic marriages could have played a part as some Yamnaya individuals were buried in Maykop kurgans and Yamnaya chieftains could have intermarried with Maykop chieftains.
1. BU2001.A0101_Beily Ugol 2_North Caucasus - R1b-Y20993
2. GW1001.A0101_Goryachevodskiy 2_North Caucasus - R1b-Z2103
3. I1723_Goryachevodskiy 2_North Caucasus - R1b-L23
4. KBD001_Kabardinka_Late North Caucasus - R1b-Z2103
5. LYG001.A0101_Lysogorskaya 6_North Caucasus - R1b-Z2103
6. MK5009.A0101_Marinskaya 5_North Caucasus - R1b-Y20993
7. KDC001.A0101_Kudachurt 3823.5_MBA North Caucasus - J2b-L283
8. PG2001_Progress 2_Eneolithic Steppe - R1b-V1636
9. PG2004_Progress 2_Eneolithic Steppe - R1b-V1636
10. 12056_Unakozovskaya 6477.4_Eneolithic Caucasus - J2a-Y11200
11. VEK007_Velikent_Kura-Araxes - J1-Z1842
12. ARM002_Kaps_Kura-Araxes - G2b
13. I1635_Kalavan_Kura-Araxes - R1b-V1636
14. RK1001.C0101_Rasshevatskiy 1_Yamnaya Caucasus - R1b-Z2103
15. RK4002.B0101_Rasshevatskiy 4_Catacomb - R1b-Z2103
16. RK4001.A0101_Rasshevatskiy 4_Catacomb - R1b-Z2108
17. SA6003.B0101_Sharakhalsun 6_Catacomb - R1b-Z2103
18. SA6013.B0101_Sharakhalsun 6_Steppe Maykop Outlier - R1a-M459(YP1272?)
19. SA6010.A0101_Sharakhalsun 6_Yamnaya Caucasus - T-L206(Y8614?)
20. I6266_Klady 5200.0_Maykop - J2a-Y11200
21. I6268_Klady 5564.0_Maykop - J2a-Y11200
22. I2051_Marchenkova Gora D13 3260.0_Dolmen LBA - J2a-M410
23. MK5008.B0101_Marinskaya 5_Late Maykop - J1(Z1842?)
24. I1720_Baksanenok_Maykop - K(L?)
25. SA6004_Sharakhalsun 6_Steppe Maykop - Q-L933
26. NV3001_Nevinnomiskiy 3_Lola - Q-L717
|
|
|
|
Post by Admin on Jul 2, 2018 18:56:11 GMT
 The first samples from the Eneolithic (one ca. 4300 BC?, the other ca. 4100 BC) are R1b1, without further subclades, so it is difficult to say if they were V88. On the PCA, they seem to be an important piece of the early Khvalynsk -> early Yamna transition period, since they cluster closer to (or even among) subsequent Yamna samples. From 3000 BC onwards, all samples from the Northern Caucasus group of Yamna are R1b-M269, which right now is probably no surprise for anyone. The Catacomb culture is dominated by R1b-Z2103, which agrees with what we saw in the unclassified Ukraine Eneolithic sample. However, the new samples (clustering close to Yamna, but with slightly ‘to the south’ of it) don’t seem to cluster closely to that first sample, so that one may still remain a real ‘outlier’, showing incoming influence (through exogamy) from the north. The admixed population from the Caucasus probably points to an isolated region of diverse peoples and languages even in this period, which justifies the strong differences among the historic language families attested in the Caucasus. So, not much space for Anatolian migrating with those expected Maykop samples with EHG ancestry, unless exogamy is proposed as a source of language change.  The Yamnaya culture from Samara is only half EHG dated to 3,300 BC. The Samara region was 100% EHG (Haak et al. 2015). So we can see in the genetic record the Caucasus influencing the steppe. |
|
|
|
Post by Admin on Sept 18, 2018 23:51:50 GMT
The later Yamnaya steppe herders of ∼3,000 BC plot between the EHG and the present-day Near East/Caucasus and the Yamnaya have ancestry from populations related to the Caucasus and South Asia that is largely absent in in Late Neolithic or Bronze Age individuals (Haak et al. 2015). This clearly shows that the Yamnaya steppe herders were a hybrid population between the EHG and populations from the present-day Near East/Caucasus, which may be closely associated with the Maikop culture. The Yamnaya culture was formed in the Pontic-Caspian steppes under the strong cultural influence of the Maikop culture with a genetic input from the Maikop people originally from the south of the the North Caucasus piedmont region or northern Mesopotamia. 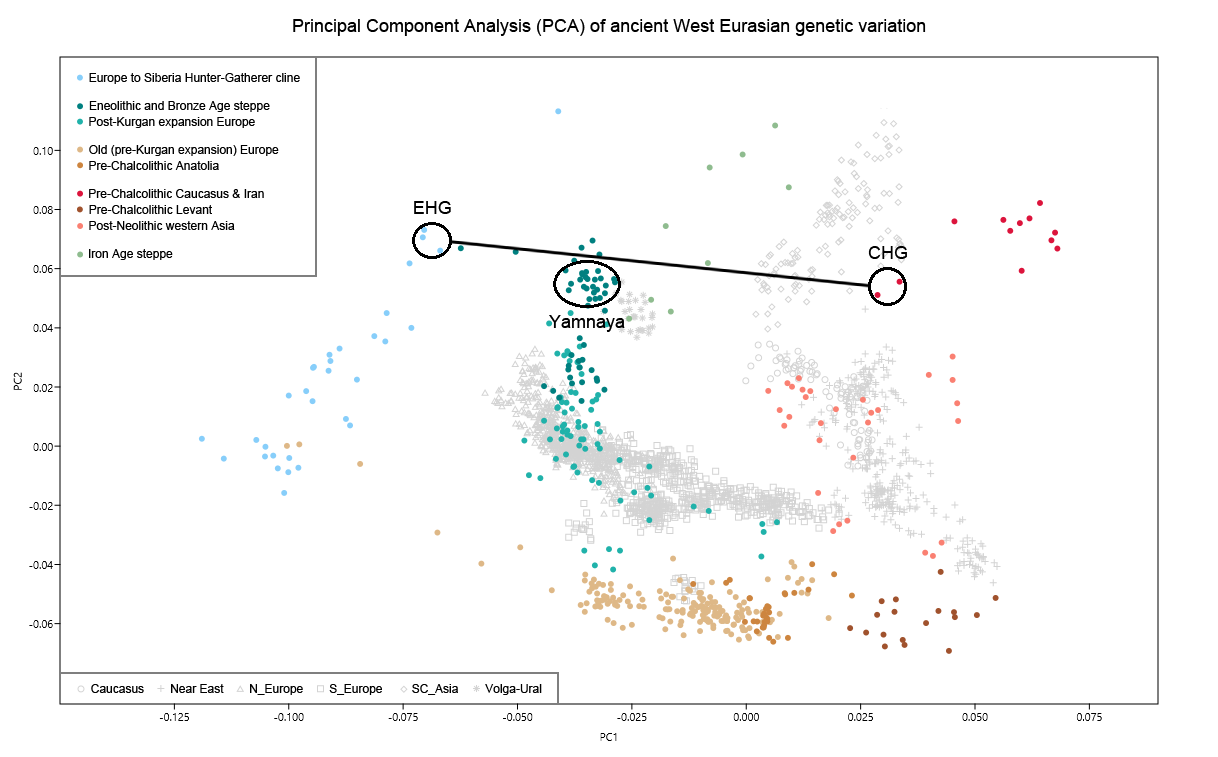 R1a and R1b are the most common haplogroups in many European populations today18,19, and our results suggest that they spread into Europe from the East after 3,000 BC. Two hunter-gatherers from Russia included in our study belonged to R1a (Karelia) and R1b (Samara), the earliest documented ancient samples of either haplogroup discovered to date. These two hunter gatherers did not belong to the derived lineages M417 within R1a and M269 within R1b that are predominant in Europeans today18,19, but all 7 Yamnaya males did belong to the M269 subclade18 of haplogroup R1b. Principal components analysis (PCA) of all ancient individuals along with 777 present-day West Eurasians4 (Fig. 2a, Supplementary Information section 5) replicates the positioning of present-day Europeans between the Near East and European hunter-gatherers4,20, and the clustering of early farmers from across Europe with present day Sardinians3,4, suggesting that farming expansions across the Mediterranean to Spain and via the Danubian route to Hungary and Germany descended from a common stock. By adding samples from later periods and additional locations, we also observe several new patterns. All samples from Russia have affinity to the ∼24,000-year-old MA1(ref. 6), the type specimen for the Ancient North Eurasians (ANE) who contributed to both Europeans4 and Native Americans4,6,8. The two hunter-gatherers from Russia (Karelia in the northwest of the country and Samara on the steppe near the Urals) form an ‘eastern European hunter-gatherer’ (EHG) cluster at one end of a hunter-gatherer cline across Europe; people of hunter-gatherer ancestry from Luxembourg, Spain, and Hungary sit at the opposite ‘western European hunter-gatherer’4 (WHG) end, while the hunter-gatherers from Sweden4,8 (SHG) are intermediate. Against this background of differentiated European hunter-gatherers and homogeneous early farmers, multiple population turnovers transpired in all parts of Europe included in our study. Middle Neolithic Europeans from Germany, Spain, Hungary, and Sweden from the period, ∼4,000–3,000 BC are intermediate between the earlier farmers and the WHG, suggesting an increase of WHG ancestry throughout much of Europe. By contrast, in Russia, the later Yamnaya steppe herders of ∼3,000 BC plot between the EHG and the present-day Near East/Caucasus, suggesting a decrease of EHG ancestry during the same time period. The Late Neolithic and Bronze Age samples from Germany and Hungary2 are distinct from the preceding Middle Neolithic and plot between them and the Yamnaya. This pattern is also seen in ADMIXTURE analysis (Fig. 2b, Supplementary Information section 6), which implies that the Yamnaya have ancestry from populations related to the Caucasus and South Asia that is largely absent in 38 Early or Middle Neolithic farmers but present in all 25 Late Neolithic or Bronze Age individuals. 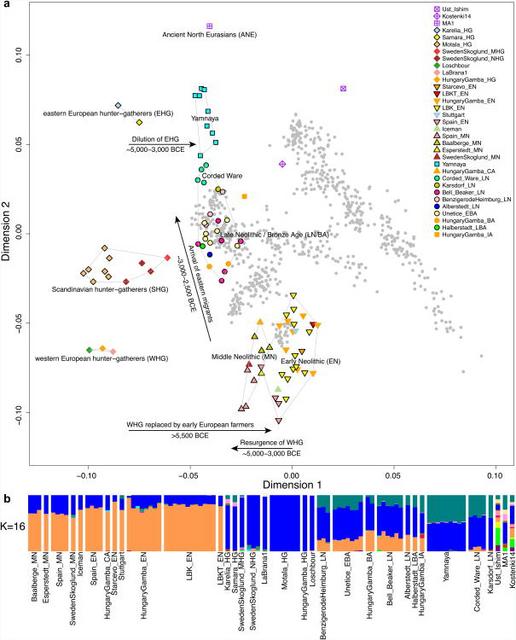 Figure 2 Population transformations in Europe (a) PCA analysis, (b) ADMIXTURE analysis. The full ADMIXTURE analysis including present-day humans is shown in Supplementary Information section 6. This ancestry appears in Central Europe for the first time in our series with the Corded Ware around 2,500 BC (Supplementary Information section 6, Fig. 2b). The Corded Ware shared elements of material culture with steppe groups such as the Yamnaya although whether this reflects movements of people has been contentious21. Our genetic data provide direct evidence of migration and suggest that it was relatively sudden. The Corded Ware are genetically closest to the Yamnaya ∼2,600km away, as inferred both from PCA and ADMIXTURE (Fig. 2) and FST (0.011±0.002) (Extended Data Table 3). If continuous gene flow from the east, rather than migration, had occurred, we would expect successive cultures in Europe to become increasingly differentiated from the Middle Neolithic, but instead, the Corded Ware are both the earliest and most strongly differentiated from the Middle Neolithic population. ‘Outgroup’ f3 statistics6 (Supplementary Information section 7),which measure shared genetic drift between a pair of populations (Extended Data Fig. 1), support the clustering of hunter-gatherers, Early/Middle Neolithic, and Late Neolithic/Bronze Age populations into different groups as in the PCA (Fig. 2a).We also analysed f4 statistics, which allow us to test whether pairs of populations are consistent with descent from common ancestral populations, and to assess significance using a normally distributed Z score. Early European farmers from the Early and Middle Neolithic were closely related but not identical. This is reflected in the fact that Loschbour, a WHG individual fromLuxembourg4, shared more alleles with post-4,000 BC European farmers from Germany, Spain, Hungary, Sweden and Italy than with early farmers of Germany, Spain, and Hungary, documenting an increase of hunter-gatherer ancestry in multiple regions of Europe during the course of the Neolithic. The two EHG form a clade with respect to all other present-day and ancient populations (|Z|<1.9), and MA1 shares more alleles with them (|Z|>4.7) than with other ancient or modern populations, suggesting that they may be a source for the ANE ancestry in present Europeans4,12,22 as they are geographically and temporally more proximate than Upper Paleolithic Siberians. The Yamnaya differ from the EHG by sharing fewer alleles with MA1 (|Z|=6.7) suggesting a dilution of ANE ancestry between 5,000–3,000 BC on the European steppe. This was likely due to admixture of EHG with a population related to present-day Near Easterners, as the most negative f3 statistic in the Yamnaya (giving unambiguous evidence of admixture) is observed when we model them as a mixture of EHG and present-day Near Eastern populations like Armenians (Z=-6.3); Supplementary Information section 7). The Late Neolithic/Bronze Age groups of central Europe share more alleles with Yamnaya than the Middle Neolithic populations do (|Z|=12.4) and more alleles with the Middle Neolithic than the Yamnaya do (|Z|=12.5), and have a negative f3 statistic with the Middle Neolithic and Yamnaya as references (Z=-20.7), indicating that they were descended from a mixture of the local European populations and new migrants from the east. Moreover, the Yamnaya share more alleles with the CordedWare (|Z|≥3.6) than with any other Late Neolithic/Early Bronze Age group with at least two individuals (Supplementary Information section 7), indicating that they had more eastern ancestry, consistent with the PCA and ADMIXTURE patterns (Fig. 2). |
|
|
|
Post by Admin on Feb 5, 2019 17:59:04 GMT
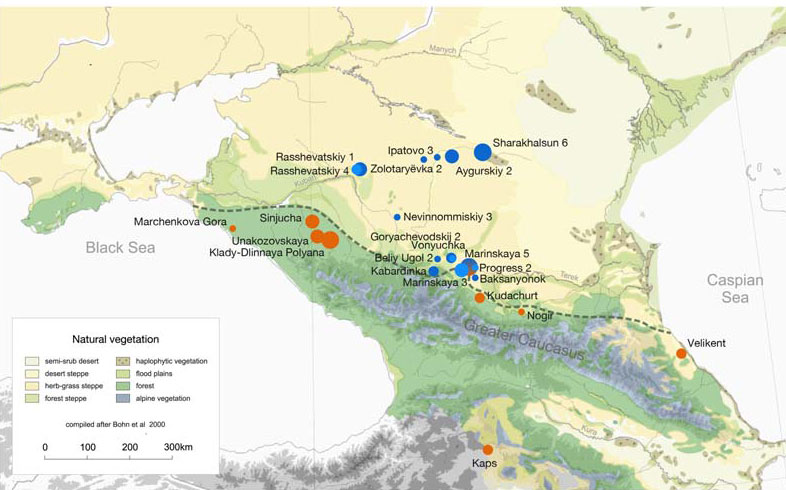
The 1100-kilometre long Caucasus mountain ranges extend between the Black Sea and the Caspian Sea and are bounded by the rivers Kuban and Terek in the north and the Kura and Araxes rivers in the south (Fig. 1). The rich archaeological record suggests extensive human occupation since the Upper Palaeolithic1,2,3. A Neolithic lifestyle based on food production began in the Caucasus after 6000 calBCE4. As a region rich in natural resources such as ores, pastures and timber, the Caucasus gained increasing importance to the economies of the growing urban centres in northern Mesopotamia5,6. In the 4th millennium BCE the archaeological record attests to the presence of the Maykop and Kura-Araxes, two major cultural complexes of the Bronze Age (BA) in the region (Fig. 1, Supplementary Note 1). The Maykop culture is well known for its large and rich burial mounds, especially at the eponymous Maykop site, which reflect the rise of a new system of social organization7, while the Kura-Araxes is found on both flanks of the Caucasus mountain range, demonstrating a connection between north and south5. 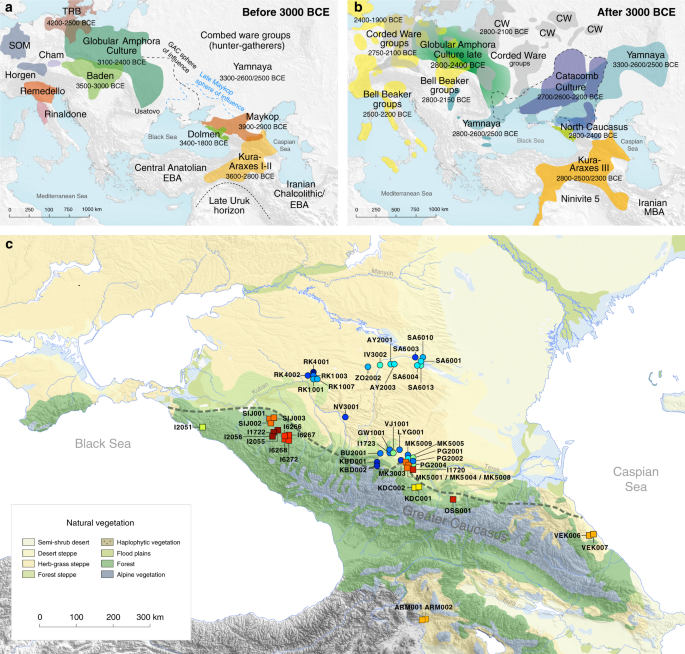 Fig. 1 Temporal and geographic distribution of archaeological cultures is shown for two windows in time a, b that are critical for our data. The zoomed map c shows the location of studied individuals from various sites in the Caucasus. Symbols and sample names correspond with Fig. 2 and Supplementary Data 1. The dashed line illustrates a hypothetical geographic border between genetically distinct Steppe and Caucasus clusters. (BB Bell Beaker; CW Corded Ware; TRB Trichterbecher/Funnel Beaker; SOM Seine-Oise-Marne complex). (All three maps were prepared by S. Reinhold and D. Mariaschk based on freely available geological and vegetation GIS-data from www.usgs.gov/, www.naturalearthdata.com/ and modified after Stone, T.A., and P. Schlesinger. 2003. RLC Vegetative Cover of the Former Soviet Union, 1990. ORNL DAAC, Oak Ridge, Tennessee, USA. doi.org/10.3334/ORNLDAAC/700.)Contact between the near East, the Caucasus, the Steppe and central Europe is documented, both archaeologically and genetically, as early as the 5th millennium BC8,9,10. This increased in the 4th millennium BCE along with the development of new technologies such as the wheel and wagon, copper alloys, new weaponry, and new breeds of domestic sheep11. Such contact was critical in the cultural12 and genetic formation of the Yamnaya complex on the Eurasian Steppe—with about half of BA Steppe ancestry thought to derive from the Caucasus13. In the 3rd millennium BC, increased mobility associated with wheeled transport and the intensification of pastoralist practices led to dramatic expansions of populations closely related to the Yamnaya14,15,16, accompanied by the domestication of horses17 allowing more efficient keeping of larger herds. These expansions ultimately contributed a substantial fraction to the ancestry of present-day Europe and South Asia18,19,20. Thus, the Caucasus region played a crucial role in the prehistory and formation of Eurasian genetic diversity. Recent ancient DNA studies have resolved several long-standing questions regarding cultural and population transformations in prehistory. One important feature is a cline of European hunter-gatherer (HG) ancestry that runs roughly from West to East (hence WHG and EHG; blue component in Fig. 2a,c). This ancestry differs from that of Early European farmers, who are more closely related to farmers of northwest Anatolia21,22 and also to pre-farming Levantine individuals9. The near East and Anatolia have long-been seen as the regions from which European farming and animal husbandry emerged. In the Mesolithic and Early Neolithic, these regions harboured three divergent populations, with Anatolian and Levantine ancestry in the west, and a group with a distinct ancestry in the east. The latter was first described in Upper Pleistocene individuals from Georgia (Caucasus hunter-gatherers; CHG)13 and then in Mesolithic and Neolithic individuals from Iran9,23. The following millennia, spanning the Neolithic to BA, saw admixture between these ancestral groups, leading to a pattern of genetic homogenization of the source populations9. North of the Caucasus, Eneolithic and BA individuals from the Samara region (5200–4000 BCE) carry an equal mixture of EHG- and CHG/Iranian ancestry, so-called ‘steppe ancestry’13 that eventually spread further west18,19, where it contributed substantially to present-day Europeans, and east to the Altai region as well as to South Asia9. 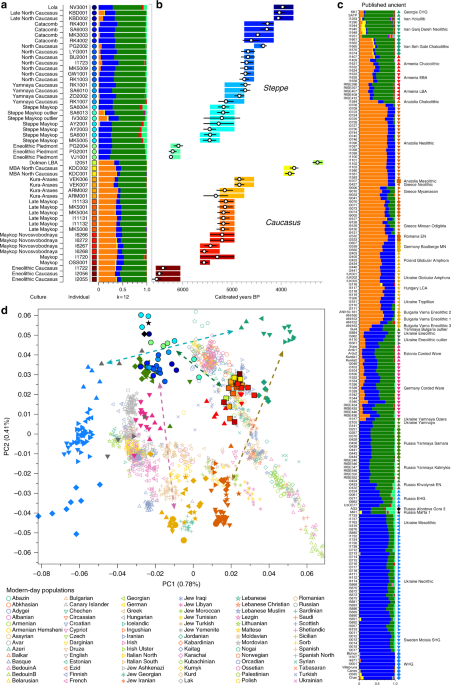 Fig. 2 ADMIXTURE and PCA results, and chronological order of ancient Caucasus individuals. a ADMIXTURE results (k = 12) of the newly genotyped individuals (filled symbols with black outlines) sorted by genetic clusters (Steppe and Caucasus) and in chronological order (coloured bars indicate the relative archaeological dates, b white circles the mean calibrated radiocarbon date and the errors bars the 2-sigma range. c ADMIXTURE results of relevant prehistoric individuals mentioned in the text (filled symbols), and d shows these projected onto a PCA of 84 modern-day West Eurasian populations (open symbols). Dashed arrows indicate trajectories of admixture: EHG—CHG (petrol), Yamnaya—Central European MN (pink), Steppe—Caucasus (green), and Iran Neolithic—Anatolian Neolithic (brown) To understand and characterize the genetic variation of Caucasian populations, present-day groups from various geographic, cultural/ethnic and linguistic backgrounds have been analyzed previously24,25,26. Yunusbayev and colleagues described the Caucasus region as an asymmetric semipermeable barrier based on a higher genetic affinity of southern Caucasus groups to Anatolian and near Eastern populations and a genetic discontinuity between these and populations of the North Caucasus and the adjacent Eurasian steppes. While autosomal and mitochondrial DNA data appear relatively homogeneous across the entire Caucasus, the Y-chromosome diversity reveals a deeper genetic structure attesting to several male founder effects, with striking correspondence to geography, ethnic and linguistic groups, and historical events24,25. In our study, we aimed to investigate when and how the genetic patterns observed today were formed and test whether they have been present since prehistoric times by generating time-stamped human genome-wide data. We were also interested in characterizing the role of the Caucasus as a conduit for gene-flow in the past and in shaping the cultural and genetic makeup of the wider region (Supplementary Note 1). This has important implications for understanding the means by which Europe, the Eurasian steppe zone, and the earliest urban centres in the Near East were connected6. We aimed to genetically characterise individuals from cultural complexes such as the Maykop and Kura-Araxes and assessing the amount of gene flow in the Caucasus during times when the exploitation of resources of the steppe environment intensified, since this was potentially triggered by the cultural and technological innovations of the Late Chalcolithic and EBA around 4000–3000 BCE5 (Supplementary Note 2). Finally, since the spread of steppe ancestry into central Europe and the eastern steppes during the early 3rd millennium BCE was a striking migratory event in human prehistory18,19, we also retraced the formation of the steppe ancestry profile and tested for influences from neighbouring farming groups to the west or early urbanization centres further south. |
|
|
|
Post by Admin on Feb 6, 2019 18:01:23 GMT
Genetic clustering and uniparentally inherited markers We report genome-wide data at a targeted set of 1.2 million single nucleotide polymorphisms (SNPs)18,27 for 59 Eneolithic and BA individuals from the Caucasus region. After filtering out 14 individuals that were first-degree relatives or showed evidence of contamination (Supplementary Data 1, Supplementary Note 3) we retained 45 individuals for downstream analyses using a cutoff of 30,000 SNPs. We merged our newly generated samples with previously published ancient and modern data (Supplementary Data 2). We first performed principal component analysis (PCA)28 and ADMIXTURE29 analysis to assess the genetic affinities of the ancient individuals qualitatively (Fig. 2). Based on PCA and ADMIXTURE plots we observe two distinct genetic clusters: one falls with previously published ancient individuals from the West Eurasian steppe (hence termed ‘Steppe’), and the second clusters with present-day southern Caucasian populations and ancient BA individuals from today’s Armenia (henceforth called ‘Caucasus’), while a few individuals take on intermediate positions between the two. The stark distinction seen in our temporal transect is also visible in the Y-chromosome haplogroup distribution, with R1/R1b1 and Q1a2 types in the Steppe and L, J, and G2 types in the Caucasus cluster (Fig. 3a, Supplementary Data 1, Supplementary Note 4). In contrast, the mitochondrial haplogroup distribution is more diverse and similar in both groups (Fig. 3b, Supplementary Data 1). 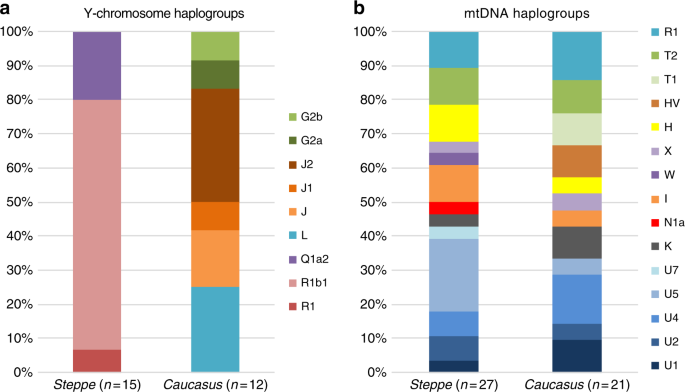 Fig. 3 The two distinct clusters are already visible in the oldest individuals of our temporal transect, dated to the Eneolithic period (~6300–6100 yBP/4300–4100 calBCE). Three individuals from the sites of Progress 2 and Vonyuchka 1 in the North Caucasus piedmont steppe (‘Eneolithic steppe’), which harbour EHG and CHG related ancestry, are genetically very similar to Eneolithic individuals from Khvalynsk II and the Samara region18,22. This extends the cline of dilution of EHG ancestry via CHG-related ancestry to sites immediately north of the Caucasus foothills (Fig. 1c; Fig. 2d). In contrast, the oldest individuals from the northern mountain flank itself, which are three first-degree-related individuals from the Unakozovskaya cave associated with the Darkveti-Meshoko Eneolithic culture (analysis label ‘Eneolithic Caucasus’) show mixed ancestry mostly derived from sources related to the Anatolian Neolithic (orange) and CHG/Iran Neolithic (green) in the ADMIXTURE plot (Fig. 2c). While similar ancestry profiles have been reported for Anatolian and Armenian Chalcolithic and BA individuals9,19, this result suggests the presence of this mixed ancestry north of the Caucasus as early as ~6500 years ago. Ancient North Eurasian ancestry in Steppe Maykop individuals Four individuals from mounds in the grass steppe zone, archaeologically associated with the ‘Steppe Maykop’ cultural complex (Supplementary Notes 1 and 2), lack the Anatolian farmer-related (AF) component when compared to contemporaneous Maykop individuals from the foothills. Instead they carry a third and fourth ancestry component that is linked deeply to Upper Paleolithic Siberians (maximized in the individual Afontova Gora 3 (AG3)30,31 and Native Americans, respectively, and in modern-day North Asians, such as North Siberian Nganasan (Supplementary Data 3). To illustrate this affinity with ‘ancient North Eurasians’ (ANE)21, we also ran PCA with 147 Eurasian (Supplementary Fig. 1A) and 29 Native American populations (Supplementary Fig. 1B). The latter represents a cline from ANE-rich steppe populations such as EHG, Eneolithic individuals, AG3 and Mal’ta 1 (MA1) to modern-day Native Americans at the opposite end. To formally test the excess of alleles shared with ANE/Native Americans we performed f4-statistics of the form f4(Mbuti, X; Steppe Maykop, Eneolithic steppe), which resulted in significantly positive Z-scores (Z >3) for AG3, MA1, EHG, Clovis and Kennewick for the ancient populations and many present-day Native American populations (Supplementary Table 1). Based on these observations we used qpWave and qpAdm methods to model the number of ancestral sources contributing to the Steppe Maykop individuals and their relative ancestry coefficients. Simple two-way models of Steppe Maykop as an admixture of Eneolithic steppe, AG3 or Kennewick do not fit (Supplementary Table 2). However, we could successfully model Steppe Maykop ancestry as being derived from populations related to all three sources (p-value 0.371 for rank 2): Eneolithic steppe (63.5 ± 2.9%), AG3 (29.6 ± 3.4%) and Kennewick (6.9 ± 1.0%) (Fig. 4; Supplementary Table 3). We note that the Kennewick related signal is most likely driven by the East Eurasian part of Native American ancestry as the f4-statistics (Steppe_Maykop, Fitted Steppe_Maykop; Outgroup1, Outgroup2) show that the Steppe Maykop individuals share more alleles not only with Karitiana but also with Han Chinese (Supplementary Table 2). 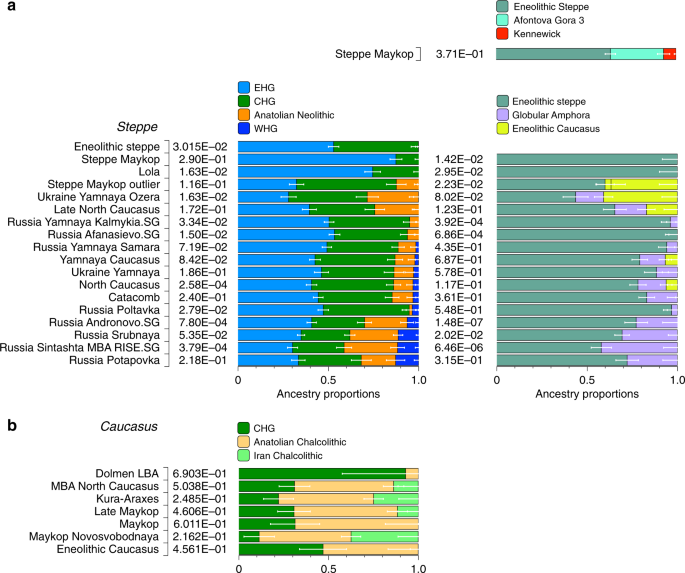 Fig. 4 Characterising the Caucasus ancestry profile The Maykop period, represented by 12 individuals from eight Maykop sites (Maykop, n = 2; a cultural variant ‘Novosvobodnaya’ from the site Klady, n = 4; and Late Maykop, n = 6) in the northern foothills appears homogeneous. These individuals closely resemble the preceding Eneolithic Caucasus individuals and present a continuation of the local genetic profile. This ancestry persists in the following centuries at least until ~3100 yBP (1100 calBCE), as revealed by individuals from Kura-Araxes from both the northeast (Velikent, Dagestan) and the South Caucasus (Kaps, Armenia), as well as MBA/LBA individuals (e.g. Kudachurt, Marchenkova Gora) from the north. Overall, this Caucasus ancestry profile falls among the ‘Armenian and Iranian Chalcolithic’ individuals and is indistinguishable from other Kura-Araxes individuals (Armenian EBA) on the PCA plot (Fig. 2), suggesting a dual origin involving Anatolian/Levantine and Iran Neolithic/CHG ancestry, with only minimal EHG/WHG contribution possibly as part of the AF ancestry9. Admixture f3-statistics of the form f3(X, Y; target) with the Caucasus cluster as target resulted in significantly negative Z scores (Z < −3) when CHG (or AG3 in Late Maykop) were used as one and Anatolian farmers as the second potential source (Supplementary Table 4). We also used qpWave to determine the number of streams of ancestry and found that a minimum of two is sufficient (Supplementary Table 5).  We then tested whether each temporal/cultural group of the Caucasus cluster could be modelled as a simple two-way admixture by exploring all possible pairs of sources in qpWave. We found support for CHG as one source and AF ancestry or a derived form such as is found in southeastern Europe as the other (Supplementary Table 6). We focused on mixture models of proximal sources (Fig. 4b) such as CHG and Anatolian Chalcolithic for all six groups of the Caucasus cluster (Eneolithic Caucasus, Maykop and Late Makyop, Maykop-Novosvobodnaya, Kura-Araxes, and Dolmen LBA), with admixture proportions on a genetic cline of 40–72% Anatolian Chalcolithic related and 28–60% CHG related (Supplementary Table 7). When we explored Romania_EN and Bulgaria_Neolithic individuals as alternative southeast European sources (30–46% and 32–49%), the CHG proportions increased to 54–70% and 51–68%, respectively. We hypothesize that alternative models, replacing the Anatolian Chalcolithic individual with yet unsampled populations from eastern Anatolia, South Caucasus or northern Mesopotamia, will likely also provide a fit to some of the tested Caucasus groups. Models with Iran Neolithic as substitute for CHG could also explain the data in a two-way admixture with the combination of Armenia Chalcolithic or Anatolia Chalcolithic as the other source. However, models replacing CHG with EHG received no support (Supplementary Table 8), indicating no strong influence for admixture from the adjacent steppe to the north. We also found no direct evidence of EHG or WHG ancestry in Caucasus groups (Supplementary Table 9), but observed that Kura-Araxes and Maykop-Novosvobodnaya individuals had likely received additional Iran Chalcolithic-related ancestry (24.9% and 37.4%, respectively; Fig. 4; Supplementary Table 10). |
|

