|
|
Post by Admin on Aug 2, 2023 23:34:19 GMT
A team of researchers with a wide variety of backgrounds from institutions in Poland, Sweden, the U.K., Czech Republic and Ukraine has learned more about the demographic history of people living in East-Central Europe during the Bronze Age by studying the genes of people living during that time. For their paper published in the journal Nature Communications, the group conducted genetic analyses of temporal bones and/or tooth remains of 91 people. 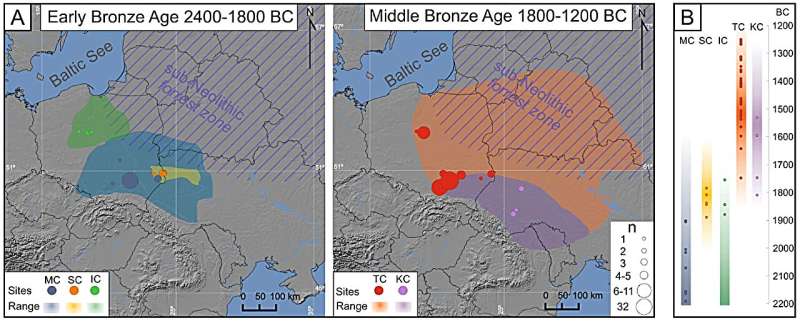 As the research team notes, most demographic research focused on early Europe has been quite generalized, spanning from the postglacial spread of hunter-gatherers to the growth of migration by farmers. In this effort, they sought to better understand such demographic events in better detail. To that end, they obtained and studied teeth and bones from people who lived in different parts of East-Central Europe during the Bronze Age. The work involved studying samples from 176 individuals, which they whittled down to 91. All the remaining samples were then sequenced, generating approximately 15,000 single-nucleotide polymorphisms, which allowed them to get a better perspective on the backgrounds of those individuals. The researchers found that most of the early people living in the region at the time most resembled the Bell Beaker and Corded Ware cultures from the Neolithic. But they also found evidence of a shift during the Bronze Age as hunter-gatherers from other parts of Europe moved in, most particularly from the northeast. The research team notes more evidence of such a shift—a move to the use of large burial plots, for example, which were similar to those often seen during the Neolithic. They describe this shift as an admixture of hunter-gatherers. The researchers also found some ancestry from an unknown group, likely people that came from Neolithic farmers. The team also found evidence suggesting that the admixture occurred further north, and those people or their offspring slowly moved south and east. They conclude that most of the people living in East-Central Europe during the Bronze Age were descendants of people from the Corded Ware culture, but also that some degree of comingling with steppe pastoralists had occurred. More information: Maciej Chyleński et al, Patrilocality and hunter-gatherer-related ancestry of populations in East-Central Europe during the Middle Bronze Age, Nature Communications (2023). DOI: 10.1038/s41467-023-40072-9 Journal information: Nature Communications www.nature.com/articles/s41467-023-40072-9 |
|
|
|
Post by Admin on Aug 11, 2023 19:18:02 GMT
Patrilocality and hunter-gatherer-related ancestry of populations in East-Central Europe during the Middle Bronze Age Abstract The demographic history of East-Central Europe after the Neolithic period remains poorly explored, despite this region being on the confluence of various ecological zones and cultural entities. Here, the descendants of societies associated with steppe pastoralists form Early Bronze Age were followed by Middle Bronze Age populations displaying unique characteristics. Particularly, the predominance of collective burials, the scale of which, was previously seen only in the Neolithic. The extent to which this re-emergence of older traditions is a result of genetic shift or social changes in the MBA is a subject of debate. Here by analysing 91 newly generated genomes from Bronze Age individuals from present Poland and Ukraine, we discovered that Middle Bronze Age populations were formed by an additional admixture event involving a population with relatively high proportions of genetic component associated with European hunter-gatherers and that their social structure was based on, primarily patrilocal, multigenerational kin-groups. Introduction Currently, it is well established that the European gene pool has been shaped by several major demographic events, including the postglacial spread of hunter-gatherers1; subsequent migrations of early farmers, which marked the beginning of the Neolithic in Europe2,3; and the later arrival of Pontic-Caspian steppe pastoralists4,5,6. However, there is still extensive debate surrounding the scale and exact nature of these demographic events and how they affected the genetic makeup of different regions across Europe7. East-Central Europe, in particular, is a region often on the frontier of these events, resulting in a mosaic of genetically distinct populations associated with a variety of cultural entities. By the turn of the Bronze Age, this region was dominated by populations associated with the Corded Ware Culture (CWC) and Bell Beaker Culture (BBC) and characterised by high levels of steppe ancestry4,5,6,8. Descendants of steppe pastoralists are thought to have replaced and admixed with the late Neolithic populations, which were characterised by large amounts of Anatolian farmer ancestry with additional low to medium levels of hunter-gatherer ancestry9,10. However, the Funnel Beaker Culture (FBC) and Globular Amphora Culture (GAC), major entities predating the arrival of steppe pastoralists associated with the later Neolithic in the region, are thought to have been long lasting, with some of their local variants continuing well into the Early Bronze Age (up to 2000 BCE, in the case of the GAC)11. As no ancient DNA (aDNA) data are available from individuals associated with later populations, their genetic makeup can only be inferred from their 3rd millennium-BC counterparts. The northern and eastern parts of East-Central Europe followed slightly different trajectories, with populations associated with the sub-Neolithic forest zone12,13, characterised by high levels of hunter-gatherer ancestry (and, to a large extent, lifestyle) throughout the Neolithic. In these regions, Anatolian farmer ancestry was introduced along with steppe ancestry during the onset of the Bronze Age14,15. Similar patterns have been observed in other regions surrounding the Baltic Sea, such as southern Scandinavia and Gotland, where hunter-gatherer individuals (both in terms of lifestyle and genetic composition) of the Pitted Ware Culture (PWC) coexisted in close proximity (but without significant gene flow) with the FBC in the 4th millennium BCE and the Battle Axe Culture (BAC), the Scandinavian variant of the CWC, in the first half of the 3rd millennium BCE16,17. The cultural landscape of Early Bronze Age (EBA) in East-Central Europe (2400–1800 BCE) is widely believed to be a direct continuation of processes that started during the onset of the epoch. For example, the cultural entities present in the region, such as those associated with the Mierzanowice, Iwno and Strzyżów archaeological cultures (MC, IC and SC, respectively)18,19,20, are largely seen as continuations of groups associated with the CWC and BBC21,22,23. In addition, steppe24,25 or the northern forest zone26 cultures have been suggested to have influenced, to some extent, the SC. The Middle Bronze Age (MBA) in the region (1800–1200 BCE) was in turn dominated by the Trzciniec Cultural Circle (TCC). This cultural phenomenon extended from the Oder River drainage basin to the Desna and Seym River basins (ca. 1200 km) and from the Baltic seashore to the Prut basin (ca. 750 km), exhibiting several territorial variants27. This study focuses on MBA individuals associated with two of these variants (Fig. 1A): the Trzciniec Culture (TC), which occupied the lands belonging to modern-day Poland and central-western Ukraine, and the Komarów Culture (KC), found in modern-day southwestern Ukraine and neighbouring parts of Romania and Moldova27. These MBA cultures retained many cultural aspects of their EBA counterparts, such as styles of pottery and bronze artefacts as well as funeral practices including under-barrow graves and cremation27. This cultural similarity have been shown to coincide with genetic continuity between CWC and both EBA and MBA populations as seen in mitochondrial genome data28. However, some elements of the TCC are unique to the EBA or MBA, particularly the predominance and scale of collective burials. Collective burials of multiple individuals were also prevalent among Middle and Late Neolithic populations in Central and East-Central Europe associated with local variants of the FBC or GAC29,30. Recent studies have shown that these Neolithic collective burials often contain remains of multiple individuals who belonged to, in most cases, patrilocal kin groups31,32,33,34. The biological relatedness among individuals found in these MBA collective burials associated with the TCC, remains unexplored. The presence or absence of kinship among these populations would, however, greatly increase our understanding of the social organization and structure of these societies. The re-emergence of collective burials is the subject of another debate concerning whether this predominant burial custom in the TCC was a result of genetic shifts or social changes within the Bronze Age populations30. Fig. 1: The geographical and temporal context and genetic affinities of the analysed Bronze Age individuals. 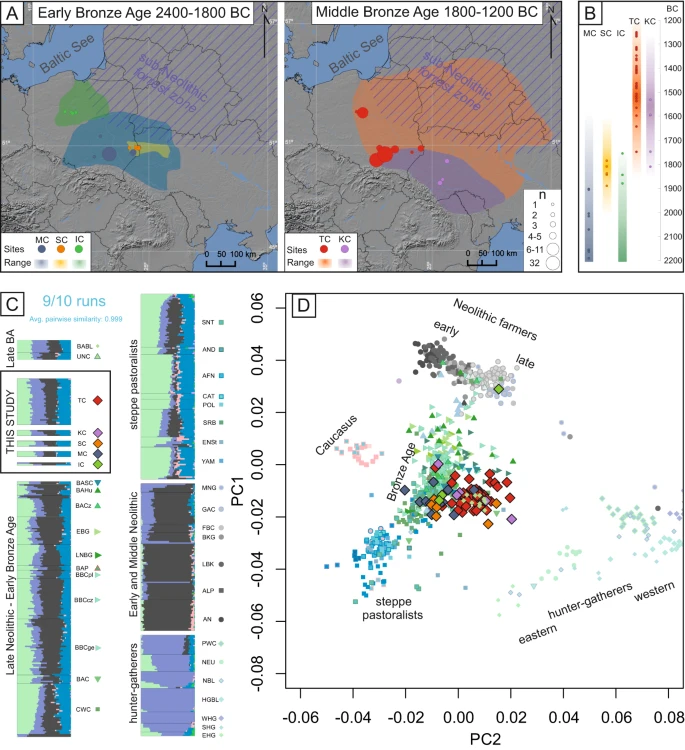 A Maps showing the locations of samples published in this study and the geographical range of their associated cultural entities; the size of the marker corresponds to the number of samples from each site. The map was created using QGIS 2.12.249 and basemap from NOAA National Geophysical Data Center. 2009: ETOPO1 1 Arc-Minute Global Relief Model. NOAA National Centers for Environmental Information. Accessed 2013. B The age of the newly generated genomes (calculated as an average of 2σ BCE dates) corresponding to the temporal range of the archaeological cultures they are associated with. C The results of unsupervised admixture analysis (K = 7) on the selected populations. D PCA plot of ancient individuals projected onto contemporary individuals from West Eurasia from the Human Origins reference panel (not shown). The symbols in both the PCA and admixture analysis correspond to individuals associated with the following cultures: IC - Iwno Culture, KC – Komarów Culture, MC – Mierzanowice Culture, SC – Strzyżów Culture, TC – Trzciniec Culture (from this study and reference populations), AFN – Afanasievo Culture, ALP – Alföld Linear Pottery Culture, AN – Anatolian Neolithic, AND – Andronovo Culture, BABL – Bronze Age Baltic, BAC – Battle Axe Culture, BACz Bronze Age Czechia, BAHU – Bronze Age Hungary, BAP – Bronze Age Poland, BASC – Bronze Age Scandinavia, BBC – Bell Beaker Culture (Poland, Czechia and Germany, respectively), BKG – Brześć Kujawski Group, CAT – Catacomb Culture, CWC – Corded Ware Culture, EBG – Early Bronze Age Germany, EHG – Eastern Hunter Gatherers, ENSt – Eneolithic Steppe, FBC – Funnel Beaker Culture, GAC – Globular Amphora Culture, HGBL – Hunter Gatherer Baltic, LBK – Linear Pottery Culture, LNBG – Late Neolithic/Bronze Age Germany, MNG – Middle Neolithic Germany, NBL – Neolithic Baltic, NEU – Neolithic Ukraine, POL – Poltavka Culture, PWC – Pitted Ware Culture, SHG – Scandinavian Hunter Gatherers, SNT – Sintashta Culture, SRB – Srubnaya Culture, UNC – Únětice Culture, WHG – Western Hunter Gatherers, YAM – Yamnaya Culture. This study, explores genetic affinities between individuals from various EBA and MBA cultures and their genetic relations to populations of preceding cultural complexes as well as possible kinship structures within MBA societies. To achieve this we conducted population genetic analyses using 91 newly generated genomes from Bronze Age individuals associated with EBA and MBA cultures from modern-day southern and south-eastern Poland and western Ukraine. |
|
|
|
Post by Admin on Aug 12, 2023 19:06:18 GMT
Results and discussion
Out of the 175 Bronze Age individuals screened, 92 produced enough data (>0.018 genome coverage) to be retained for further analysis and/or deeper sequencing. An additional 100 libraries, including 37 uracil-DNA-glycosylase (UDG)-treated libraries, were created and sequenced for the selected individuals. With the exception of two libraries from two different individuals, all libraries displayed characteristic post-mortem damage at the 5′ and 3′ ends of aDNA fragments and low contamination levels, as estimated by mitochondrial sequences and X chromosome sequences in males, where sufficient coverage was obtained (Supplementary Data 1 and Supplementary Data 2). Only one library for individual poz751, which displayed very low levels of post-mortem damage, and one of two libraries for individual poz664, in which high levels of mitochondrial DNA (mtDNA) contamination were detected, were excluded from further analysis. Therefore, the final dataset used for the kinship and population genetic analyses consisted of 91 individuals with a median genome coverage of 0.2× (ranging from 0.019× to 2.29×), representing the IC (n = 3), MC (n = 15), SC (n = 6), TC (n = 62) and KC (n = 5) cultures.
Genetic affinities of Early Bronze Age populations from East-Central Europe indicate the continuation of processes initiated in the onset of the epoch
The majority of the EBA individuals in this study (2200–1850 BCE) associated with the MC, IC and SC are genetically similar to their direct cultural predecessors (such as the BBC and CWC), as indicated by the principal component analysis (PCA) plot (Fig. 1D). The results of unsupervised admixture analysis (K = 7) of the selected populations (Fig. 1C) support these relationships as they indicate similar levels of admixture components (Fig. 1C). These findings are consistent with the general archaeological consensus and previous analyses of mitochondrial data28 Although the IC is believed to have the greatest cultural similarity to groups associated with the BBC, the one individual analysed here (poz929) exhibited closer affinity with individuals associated with various CWC groups rather than the BBC populations according to the f3 and D statistics (Supplementary Data 7, Supplementary Data 12 and Supplementary Fig. 2). A similar trend was observed in the case of individuals attributed to the MC, which displayed closer affinity to CWC individuals from Estonia over other EBA groups. However, the division between the BBC and CWC (and their definition as independent cultural entities associated with distinct populations) is contested7, and the majority of the above D statistics have low Z scores. Further studies are needed to fully understand the regional, cultural and genetic complexity of Central and Eastern Europe after the arrival of steppe pastoralists.
One IC-associated male from Łojewo (poz502) deviated from the general pattern, as he was genetically closest to the Middle and Late Neolithic populations, which occupied the same space on the PCA plot and displayed similar admixture proportions. The results of f3 statistics indicate that the population sharing the most genetic drift with this individual was the GAC, followed closely by the FBC (Supplementary Data 7). Such seemingly Neolithic individuals have occasionally been observed in populations postdating the arrival of steppe pastoralists and have been hypothesised to be foreigners who were incorporated into Bronze Age societies from isolated populations that retained a Middle Neolithic genetic makeup up to the end of the 3rd millennium BCE35. If this interpretation is applied to poz502, radiocarbon dated to the border between the EBA and MBA (2008-1750 BCE), it might indicate that such isolated populations lasted far longer than previously reported. This hypothesis is supported by the archaeological record, which shows that some Neolithic cultures, most notably the GAC, lasted well into the Bronze Age21.
Similar to poz502, two of the SC-associated males (poz794 and poz758) differed genetically from other EBA individuals. These two males were closer to the hunter-gatherer space in the PCA plot (Fig. 1D) and showed an increased proportion of genetic components that were maximised in various European hunter-gatherer populations in the admixture analysis (Fig. 1C). However, direct radiocarbon dating for one of these individuals (poz794, 1921–1697 BCE) as well as their genetic similarity to the MBA populations analysed in this study indicate that these two males should be discussed as a part of the genetic shifts observed in the MBA. Notably, the SC is generally thought to be a regional cultural phenomenon with mixed cultural traits, leading to frequent dispute over the association of individual burials or sites with this culture24,25,26. Therefore, the definition and associations of the SC with any genetically distinct population warrants further exploration targeting a broader selection of individuals attributed to this culture.
|
|
|
|
Post by Admin on Aug 13, 2023 18:44:32 GMT
Increase in hunter-gatherer ancestry in East-Central Europe in the Middle Bronze Age
The MBA individuals analysed here were dated to a range between 1750 and 1200 BCE and were associated with the TCC, representing both the TC and KC. The majority of these individuals clustered together in PCA space and shared similar admixture proportions (Fig. 1C and D). This apparent genetic relation is further highlighted by f3 and D statistics, which indicate that when analysed separately, KC and TC individuals do not, in majority of cases, display any statistically significant closer genetic affinity to either of the two populations (Supplementary Data 7 and Supplementary Data 12, Supplementary Figs. 3C and 4B). These results are in accordance with the archaeological interpretation that questions the separation of the TC and KC, arguing in favour of treating them as regional variants of the same phenomenon27,36.
Interestingly compared to EBA populations, the MBA individuals were closer in the PCA space to various hunter-gatherer populations from Europe (Fig. 1D), something that previously was not detected in analyses of mitochondrial genome data alone28. Moreover, admixture analysis indicated elevated amounts of genetic components maximised in hunter-gatherers (Fig. 1C). This suggests an additional admixture event at the beginning of the MBA involving a population with relatively high proportions of this genetic component. However, there were notable deviations to this trend, with three individuals associated with TC from Pielgrzymowice site and poz643, a relatively early KC male from Beremiany, clustering closer to EBA populations in PCA space and displaying the lowest levels of shared genetic drift with both TC and hunter-gatherer populations (Supplementary Data 7). When using qpAdm to test for possible two-way admixture models that resulted in the formation of MBA populations, several models were determined to be plausible (Supplementary Data 13) with the highest p value (p = 0.21) obtained for pair consisting of IC and Neolithic Baltic hunter-gatherers (NBL). Similarly high pvalues were found for other pairs including IC and other hunter-gatherer populations: Western Hunter Gatherers (WHG), PWC hunter-gatherers from Gotland, hunter-gatherer buried in BKG context (BKGout) and hunter-gatherer populations predating the NBL (HGBL) (p = 0.207, 0.204, 0.164, 0.160 respectively). These results are in accordance with the archaeological hypothesis that the IC greatly contributed to the emergence of the TC27. However, this finding should be interpreted with caution, as only one individual associated with the IC had sufficient coverage for inclusion in qpAdm. High fit values were also obtained for pair of populations, containing individuals from modern day Estonia associated with the CWC, as the EBA predecessor and PWC, as the source of hunter-gatherer ancestry (p = 0.11).
The process of admixture was likely more complex, as indicated by the three-way admixture model with the addition of a Neolithic population. These models yielded even better fits than the two-way models. Multiple plausible scenarios were found, all displaying high fit values (p > 0.9) including CWCes in addition to various hunter-gatherer and Neolithic populations (Supplementary Data 14). The more discriminatory rotating outgroup approach to qpAdm37 used for both two- and three- way models helped to narrow down the number of plausible scenarios. After excluding the models containing sources consisting of one individual, only three-way scenarios including CWCes and either GAC or FBC as Neolithic and NBL or HGBL as hunter-gatherer population were found to be plausible (Supplementary Data 15 and Supplementary Data 16). Based on geographical and temporal proximity as well as results of D statistics directly comparing the potential ancestry sources for each MBA individual (Supplementary Data 12, Supplementary Fig. 3) we find CWCes, NBL and GAC to be the best proxies for populations involved in the admixture process. However high fit values and close genetic affinity to the individual with hunter-gatherer ancestry buried in BKG context, indicate that further studies might help to better define the populations involved in the process.
The most likely hypothesis is that these admixed MBA populations originated in the confluence of the sub-Neolithic forest zone, associated with populations with dominant WHG ancestry14,15 as well as post-CWC groups characterised by a large proportion of steppe ancestry. The sub-Neolithic forest zone” is a broad term that includes various archaeological cultures from north-eastern Europe, characterised by long-lasting preservation of a predominantly hunter-gatherer lifestyle, and the incorporation of cultural elements of Neolithic and Bronze Age origin38. These populations remained genetically distinct from the Neolithic and post-Neolithic populations, although they maintained some level of long lasting cultural and economic exchange14 It is possible that this lead to some degree of gene flow between those populations, similar to the one observed in the case of PWC in Gotland6,16 followed by subsequent contacts with EBA descendants of steppe pastoralists. Moreover, the TCC and sub-Neolithic forest zone exhibited similar cultural traits, mostly in the form of pottery and technologies12,13,39,40,41. These similarities have often been interpreted as signs of primarily cultural exchange. Our results, showing an increase in WHG ancestry during the MBA, indicate that at least some level of admixture occurred during these interactions.
Notably, the two EBA individuals (poz794 and poz758) associated with the SC, that displayed closer genetic affinity to the MBA populations both came from the south-eastern part of modern-day Poland (Supplementary Text); of these two, the individual with a direct date (poz794) predated the MBA samples analysed here. This observation suggests that the contact zone described above is not the only place where admixture took place and/or that the process was more geographically diffused. Either is plausible, given the range and duration of exchange networks seen in the EBA40. Individual poz794 might even signal the beginning of the observed gene flow, which would date it to approximately 1800 BCE.
The SC is usually seen as a continuation of CWC traditions with additional elements from steppe cultures such as the Catacomb Culture24,25. However, the genetic shift toward an increase in WHG ancestry cannot be explained by additional migration from the steppe, a notion that we previously proposed based solely on mitochondrial data28. This idea is not supported by any indication of increase of steppe ancestry as calculated by three-way qpAdm modelling including WHG, AN and YAM as best proxies of major ancestries in European gene pool (Supplementary Data 17, Supplementary Fig. 4A). Moreover, two-way qpAdm models that explored the scenarios resulting in the emergence of the MBA populations, including EBA individuals and hunter-gatherer populations, yielded higher probabilities than models including additional steppe populations such as Andronovo, Afanasievo, Sintashta, Poltavka, Karasuk, or Srubnaya (Supplementary Data 13).
The process of admixture, which began around 1800 BCE, appears to have been a continuous rather than a result of a single migratory event, as evidenced by the presence of individuals with very high or very low proportions of hunter-gatherer ancestry throughout the whole temporal range of MBA samples analysed here (Fig. 2C, Supplementary Data 17). However, gene flow was likely more extensive in the beginning, as both shared genetic drift (as identified by f3 statistics) and admixture proportions (calculated with qpAdm) show that the proportion of hunter-gatherer ancestry decreased slightly over time (Fig. 2A and Fig. 2C). The results of this event must have been long lasting, as Late Bronze Age individuals from modern-day Latvia and Lithuania14 retain the same genetic composition as our MBA individual despite living nearly half a century later and were found, based on f3 statistics, to have the closest genetic affinity to the MBA individuals presented here out of all Bronze Age populations (Supplementary Data 7 and Supplementary Fig. 3C).
|
|
|
|
Post by Admin on Aug 14, 2023 17:53:11 GMT
Fig. 2: The hunter-gatherer ancestry in the Middle Bronze Age populations from East-Central Europe. 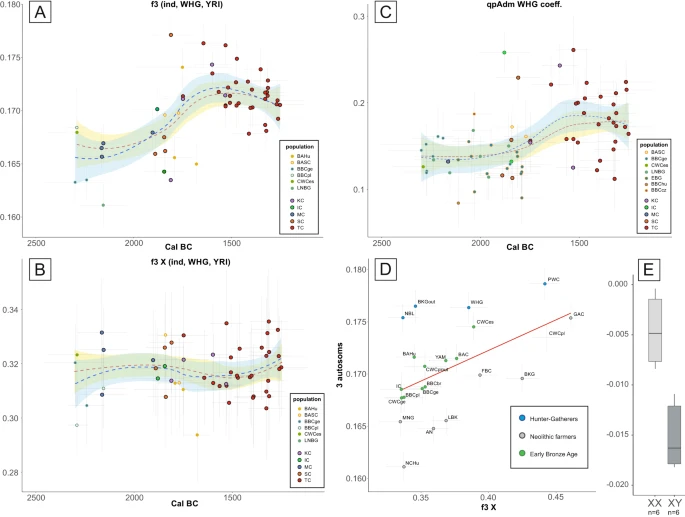 A The shared genetic drift between the newly published individuals and WHG hunter-gathers was estimated with the use of the f3-statistic, shown separately for autosomal and (B) X-chromosome data. C the WHG ancestry estimated for new (outlined in black) and reference individuals from the Final Neolithic to the Middle Bronze Age. The degree of ancestry was estimated from three-way admixture models including the WHG, AN and YAM, the points represent coefficient for WHG ancestry calculated using qpAdm (only values with p value for nested models <0.05). Error bars in (A–C) correspond to one standard error for the f3-statistics or qpAdm values (vertical) and 2σ for the dates (horizontal). The fit lines in (A–C) display smoothed conditional means for all individual (blue) and after removal of outliers (red) with corresponding 95% confidence intervals (light blue and yellow respectively). D Outgroup f3 statistics values of form f3_Xchr(YRI, TC, popX) and f3_Autosomes(YRI, TC, popX) plotted against each other with error bars representing one standard deviation for each value, red line represents linear regression inserted for visualisation purpose. E The patrilocal character of Żerniki Górne cemetery as shown by the difference in the D-statistic in the form D(YRI, Żerniki individual; Żerniki, TC) for the two sexes. The boxplots show the median (middle horizontal line), interquartile range (25th and 75th percentile) indicated with lower (25th percentile) and upper (75th percentile) hinges of the box, and whiskers extending to the lowest (highest) value that is within 1.5 times the interquartile range of the upper (lower) hinge. The labels in all panels are as follows: IC – Iwno Culture, KC – Komarów Culture, MC – Mierzanowice Culture, SC – Strzyżów Culture, TC – Trzciniec Culture, AN – Anatolia Neolithic, BAC – Battle Axe Culture, BAHu – Bronze Age Hungary, BAP – Bronze Age Poland, BASC – Bronze Age Scandinavia, BBC – Bell Beaker Culture, BKG – Brzesc Kujawski Group, CWC – Corded Ware Culture, EBG – Early Bronze Age Germany, FBC – Funnel Beaker Culture, GAC – Globular Amphora Culture, LNBG – Late Neolithic/Bronze Age Germany, MNG Middle Neolithic Germany, NBL Neolithic Baltic, NCHu Neolithic/Chalcolithic Hungary. Furthermore, several lines of reasoning support the idea that this admixture event was dominated by males originating in a population characterised by a high level of hunter-gatherer ancestry. First, as shown by our direct kinship analyses below, the resulting population was primarily patrilocal. Second, the MBA composition of Y-DNA haplogroups differed significantly from the predating populations, as dominance of I2a1a and I2a1b haplogroups was previously seen only sporadically in various hunter-gatherer populations, including two Narva Culture individuals14 and interestingly in high frequency, although for different sub-haplogroups, in GAC collective burials10,34,42. The I2a1 haplogroups were found in 75% of TC-associated MBA individuals, even after selecting only one individual from each detected kin group. Moreover, this shift was not apparent when looking at the mitochondrial haplogroups28. Finally, direct analysis of genetic distances in X-chromosome data, as determined by f3 statistics using the approach suggested by Saag et al.15 showed that on autosomes TC was relatively more similar to hunter-gatherer populations than on X chromosome (Fig. 2D, Supplementary Data 5 and Supplementary Data 6). In addition, when looking at the temporal changes in X-based f3 values we did not observe increased amounts of genetic drift shared with WHG individuals between 1800 and 1500 BCE, as seen in the autosomal data (Fig. 2B, Supplementary Data 8). The exact trajectory of events leading to the genetic shift in the MBA cannot be reconstructed with current knowledge. The Eastern Baltic hunter-gatherer populations were associated with multiple archaeological cultures that engaged in direct contact with Neolithic farmers for millennia. It cannot be excluded that at some points of this coexistence, migratory events occurred, leading to the emergence of admixed populations that, in turn, later mixed with steppe pastoralists or their Central European descendants, resulting in the formation of MBA populations analysed here. The lack of more diverse genetic data from East-Central Europe prevents us from pinpointing the exact populations that took part in this admixture. As the archaeological record shows that contact between culturally distinct groups of farmers and hunter-gatherers were long lasting, leading to substantial cultural changes38, it is possible that the practices of collective burials and patrilocal residence were some of those changes. This could be reflected in the high frequency of I2a1b Y haplogroup in some collective burials associated with middle Neolithic GAC culture10,34,42.The observed changes could have resulted from several processes involving multiple populations; our observations represent the sum of those processes. |
|

