Post by Admin on Nov 14, 2020 23:44:02 GMT
Diagenesis and the completeness of the archaeological record
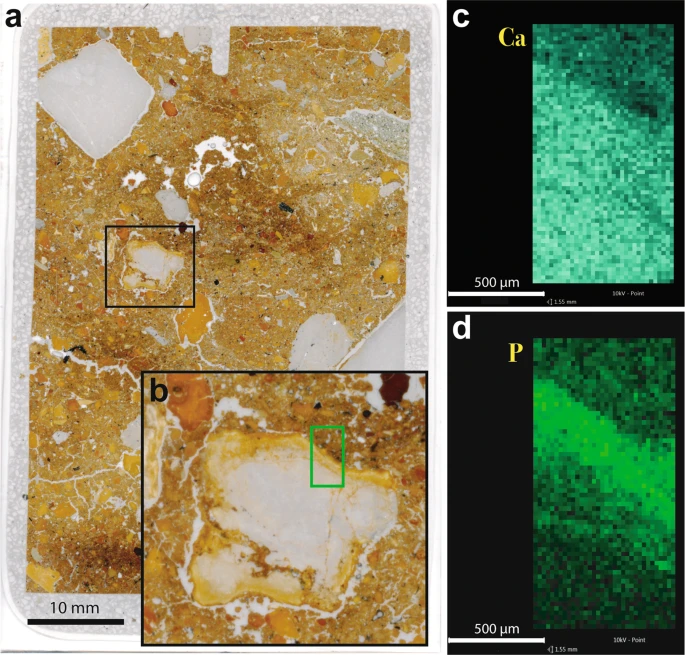
Chemical alteration features are rare in Denisova Cave. Where present, they take the form of carbonate dissolution and phosphatisation, such as that reported for the uppermost Pleistocene and Holocene layers in DCE38. In thin section, we observe phosphatic rinds around limestone clasts, a common occurrence in prehistoric caves when calcite reacts to phosphate-rich solutions39,40,41,42,43,44. This is expressed as reaction rims around individual clasts (Fig. 5), resulting in replacement of the original birefringent calcite by isotropic phosphate, generally apatite (dahllite). We also record the etching of calcite sand and decalcification of the surrounding matrix (e.g., in layer 13 in DCE), indicating the dissolution of calcium carbonate.
Figure 5
Animals are the most likely source of phosphate in an archaeological cave sequence40. Although some layers are richer than others in phosphates, including coprolites, none of those examined in thin section stands out as being excessively phosphatised. Bat and bird guano is also a possible source of cave phosphates and associated diagenetic transformations21,38,40,41,42,43,44,45,46,47,48,49,50,51. We did not record guano directly in thin section, but acidic water percolating through guano—in combination with coprolite-rich sediments—can dissolve calcite21, including the fine calcareous fraction of limestone grains and the outer surface of larger limestone fragments, to produce apatite (dahllite) rims. Bat remains occur in relatively high numbers in some layers9,35. Although bats do not commonly occupy caves at the same time as hominins, small populations could, nonetheless, have supplied a persistent supply of guano to maintain phosphatisation processes.
Common diagenetic cave minerals (e.g., taranakite, leucophosphite, crandallite, brushite and ardealite)48,52,53,54,55 have been recorded in the Holocene deposits and in layer 11.1 in DCE38. The diagenesis occurred during the Holocene and affected only the upper parts of the underlying Pleistocene sediments. We do not observe these minerals in our thin sections of the Pleistocene deposits, underscoring Denisova Cave as a depositional environment where persistently cold conditions have afforded exceptional preservation of organic materials—including lipid micro-residues on Middle Palaeolithic stone tools in DCE56—and minimal diagenesis.
Cold-climate indicators and implications for cave use
We record platy microstructures in thin section for layers 12.2 (70 ± 8 to 58 ± 6 ka) and 11.4/11.2 (44 ± 5 to 38 ± 3 ka) in DCM, and for layers 13 (156 ± 15 to 146 ± 11 ka) and 11.1/9.1 (49 ± 8 to after 38 ± 9 ka) in DCE. These features, together with the presence of rounded grains and granostriated b-fabrics, which are indicative of grain rotation, indicate incipient cryoturbation. This modification of the sediment structure is most likely associated with seasonal frost, with the thinner bands in layer 9.1 of DCE possibly associated with repeated ice lensing as a result of soil creep during thaw57. The limestone clasts in these parts of the stratigraphy are generally angular and fresh, and lack signs of phosphatisation that would reflect diagenetic transformations of calcite. We therefore correlate these platy structures with the occurrence of low temperatures in the cave and relatively few freeze-thaw cycles58.
In DCM, these platy microstructures are associated with sediments that contain unequivocal signs of hominin occupation (charcoal and closely associated bone fragments; Fig. 6). We do not know the vertical extent of these post-depositional features, however, so it is not clear how these signatures correlate. They may penetrate down into the underlying, older layers, but the sediments immediately above and adjacent to these samples are not affected in this way. Given the slow rate of sedimentation in the cave, we cannot rule out later over-printing of the sediments by these cold-climate indicators.
Figure 6

At the present day, thin (mm-thick) vertical cracks filled with ice have sometimes been observed within the Holocene deposits in the South Chamber. These would not, however, account for the horizontal ice lensing observed in our thin sections of layers 11.2 and 11.4 in DCM, and we see no such analogous vertical fissures in any of our thin sections. It is not clear why such platy structures and signs of incipient cryoturbation are not more common in the Pleistocene sequences at Denisova Cave, but this may relate to the enclosed cave environment mitigating extremes in temperature through restricted airflow. The South Chamber is better ventilated than are DCM and DCE, which may explain their modern occurrence there.
Discussion
Micromorphological analysis of the Denisova Cave sequence has provided micro-contextualised insights into the use of the site by hominins and other animals. These new data largely support previous interpretations based on field observations and other proxy datasets (e.g., faunal and pollen records6,7,8,9,35,59), thus increasing confidence in environmental reconstructions for the cave and surrounding region.
Microscopic evidence for hominin use of the cave is minimal. Decades of excavation have generated significant numbers of stone artefacts6,9 that accumulated over a substantial time interval and represent multiple occupational pulses. Micro-remains of a hominin presence—such as combustion bi-products—are readily mobilised and re-deposited, so the lack of intact features indicating fire-use in the Pleistocene sequence is intriguing. Given the limited spatial area that our micromorphological study encompasses, this outcome could be due to sampling bias (e.g., ref.60). On the other hand, because easily dispersed combustion bi-products are, nonetheless, very rare, the early occupants of the site may not have been prolific pyrotechnologists. Where we do record charcoal, it is usually well preserved, so we can rule out the possibility of complete degradation of this material and its preferential removal from the sedimentary record.
The abundant coprolite record shows that the cave was occupied by animals near-continuously. Cave hyena (Crocuta crocuta speleans)—the dominant carnivore in the Altai during the Pleistocene6,59—was present throughout the period of deposition of the Pleistocene sediments7,9,35. Whereas bones can accumulate at a cave site through the agency of various animals, animal droppings are most likely to be delivered directly to the cave floor. Coprolites, then, can be viewed as authigenic components of the sedimentary fill and we surmise that animals—mainly carnivores—used the sampled area of the cave throughout the time represented by the preserved sediments. Fossils of cave hyena are considerably more frequent in the DCE faunal record than are those of other Pleistocene predators, such as wolf, so hyenas are most likely the main accumulating agents of the faunal remains, given the dominance of their coprolites in the cave sediments.
Coprolite fragments occur in high frequencies in layers that have been affected by frost action (e.g., layers 11.4 and 11.2 in DCE). We cannot rule out the possibility that the sampled areas fall within specific latrine areas used for ‘social defecation’61,62, perhaps exacerbated by colder temperatures driving the animals into the further recesses of the cave. At Zhoukoudian Cave in northeastern China60, rich coprolite concentrations and signs of trampling were recognised from sampling localities close to the walls of the cave, confirming the attraction of these animals to marginal zones. Profiles sampled close to the walls of caves may, therefore, fall in areas favoured for animal latrines, underscoring the importance of sampling at multiple locations throughout a site.
An important outcome of our study is the identification of microstratigraphic features consistent with freezing conditions. At present, even on a sunny summer’s day, the cave interior is cold, especially so in DCE, due to the high thermal mass of the surrounding rock mitigating the warm temperatures experienced outside the cave. The cave sediments are frozen during the winter months, when temperatures can drop to an average of −16 °C in January9, but apparently not to the extent that the sediment fabric is re-arranged by ice lensing. The platy structures developed in layers deposited during late Marine Isotope Stage (MIS) 4 and MIS 3 in DCM, and during MIS 6 and MIS 3 in DCE, suggest colder conditions than those experienced in the current interglacial. We postulate that the platy features observed do not necessarily reflect the coldest conditions at the site during the Pleistocene, but are associated with specific formation environments—cold and humid conditions—that allow ice lensing to occur63.
While we observe incipient platy structures in sediments dated to terminal MIS 3 and early MIS 2 (e.g., layer 9, deposited between about 40 and 20 ka), and thus overlapping with the coldest period of the last glacial cycle, the platiness is best expressed in layers that accumulated during the preceding, comparatively warm MIS 3. There is debate surrounding the synchronicity of glacial expansion in the Altai with global climate proxies64,65, potentially associated with increased moisture availability in central Asia during MIS 3 relative to MIS 2, which was drier. The Denisova Cave microstratigraphic record may, therefore, document both cold and relatively humid conditions66, sufficient to develop platy microstructures in the near-surface, cave floor sediments.
Our study provides spatially resolved information on the depositional and post-depositional environments that have preserved organic materials in Denisova Cave. Throughout the sequence, we record only limited evidence of severe chemical diagenesis in the Pleistocene layers, but these include layers that have also yielded sedimentary aDNA10. Even in these layers, however, the evidence for diagenesis is generally weak, albeit spatially variable, providing opportunities for DNA preservation. Cold conditions may also promote the survival of genetic material and lipids in coprolites, from which aDNA and faecal biomarkers can be extracted directly for species identification and dietary reconstruction67,68.
Conclusions
The deposits in Denisova Cave contain microscopic traces of hominin and animal activities that illuminate the use of the cave over the last three glacial–interglacial cycles (Fig. 3). The micromorphological results show that the cave sediments are predominantly geogenic (naturally occurring), augmented by biogenic (biological) additions (e.g., coprolites, guano and digested bone) and anthropogenic inputs (e.g., charcoal, stone artefacts and associated debitage).
Relationships between the various lines of evidence (e.g., micro-charcoal, bioturbation, coprolites and diagenesis), examined at a finely resolved spatial scale, reveal that hominin activities in the microstratigraphic record are few. On the other hand, coprolitic evidence for cave-dwelling carnivores is ubiquitous and suggests that the site often served as a den for hyenas and, to a lesser extent, for wolves.
The cave was visited sporadically by hominins, who appeared not to have been prolific users of fire, at least in the Middle Palaeolithic deposits that constitute the majority of the Pleistocene sequence. The low frequency of hominin occupation has implications for determining the potential agency by which the few Denisovan and Neanderthal fossils were introduced to the site and their post-depositional stratigraphic integrity9,12.
The environmental conditions that best preserve organic molecules, such as DNA and lipids, and whether these materials can be recovered from specific components of the microstratigraphy (e.g., coprolite fragments), also warrant further investigation. Ongoing work at Denisova Cave aims to more fully integrate micromorphology and sedimentary aDNA analyses to develop a predictive tool for organic material preservation in the deposits at this unique hominin locality.
Chemical alteration features are rare in Denisova Cave. Where present, they take the form of carbonate dissolution and phosphatisation, such as that reported for the uppermost Pleistocene and Holocene layers in DCE38. In thin section, we observe phosphatic rinds around limestone clasts, a common occurrence in prehistoric caves when calcite reacts to phosphate-rich solutions39,40,41,42,43,44. This is expressed as reaction rims around individual clasts (Fig. 5), resulting in replacement of the original birefringent calcite by isotropic phosphate, generally apatite (dahllite). We also record the etching of calcite sand and decalcification of the surrounding matrix (e.g., in layer 13 in DCE), indicating the dissolution of calcium carbonate.
Figure 5
Animals are the most likely source of phosphate in an archaeological cave sequence40. Although some layers are richer than others in phosphates, including coprolites, none of those examined in thin section stands out as being excessively phosphatised. Bat and bird guano is also a possible source of cave phosphates and associated diagenetic transformations21,38,40,41,42,43,44,45,46,47,48,49,50,51. We did not record guano directly in thin section, but acidic water percolating through guano—in combination with coprolite-rich sediments—can dissolve calcite21, including the fine calcareous fraction of limestone grains and the outer surface of larger limestone fragments, to produce apatite (dahllite) rims. Bat remains occur in relatively high numbers in some layers9,35. Although bats do not commonly occupy caves at the same time as hominins, small populations could, nonetheless, have supplied a persistent supply of guano to maintain phosphatisation processes.
Common diagenetic cave minerals (e.g., taranakite, leucophosphite, crandallite, brushite and ardealite)48,52,53,54,55 have been recorded in the Holocene deposits and in layer 11.1 in DCE38. The diagenesis occurred during the Holocene and affected only the upper parts of the underlying Pleistocene sediments. We do not observe these minerals in our thin sections of the Pleistocene deposits, underscoring Denisova Cave as a depositional environment where persistently cold conditions have afforded exceptional preservation of organic materials—including lipid micro-residues on Middle Palaeolithic stone tools in DCE56—and minimal diagenesis.
Cold-climate indicators and implications for cave use
We record platy microstructures in thin section for layers 12.2 (70 ± 8 to 58 ± 6 ka) and 11.4/11.2 (44 ± 5 to 38 ± 3 ka) in DCM, and for layers 13 (156 ± 15 to 146 ± 11 ka) and 11.1/9.1 (49 ± 8 to after 38 ± 9 ka) in DCE. These features, together with the presence of rounded grains and granostriated b-fabrics, which are indicative of grain rotation, indicate incipient cryoturbation. This modification of the sediment structure is most likely associated with seasonal frost, with the thinner bands in layer 9.1 of DCE possibly associated with repeated ice lensing as a result of soil creep during thaw57. The limestone clasts in these parts of the stratigraphy are generally angular and fresh, and lack signs of phosphatisation that would reflect diagenetic transformations of calcite. We therefore correlate these platy structures with the occurrence of low temperatures in the cave and relatively few freeze-thaw cycles58.
In DCM, these platy microstructures are associated with sediments that contain unequivocal signs of hominin occupation (charcoal and closely associated bone fragments; Fig. 6). We do not know the vertical extent of these post-depositional features, however, so it is not clear how these signatures correlate. They may penetrate down into the underlying, older layers, but the sediments immediately above and adjacent to these samples are not affected in this way. Given the slow rate of sedimentation in the cave, we cannot rule out later over-printing of the sediments by these cold-climate indicators.
Figure 6
At the present day, thin (mm-thick) vertical cracks filled with ice have sometimes been observed within the Holocene deposits in the South Chamber. These would not, however, account for the horizontal ice lensing observed in our thin sections of layers 11.2 and 11.4 in DCM, and we see no such analogous vertical fissures in any of our thin sections. It is not clear why such platy structures and signs of incipient cryoturbation are not more common in the Pleistocene sequences at Denisova Cave, but this may relate to the enclosed cave environment mitigating extremes in temperature through restricted airflow. The South Chamber is better ventilated than are DCM and DCE, which may explain their modern occurrence there.
Discussion
Micromorphological analysis of the Denisova Cave sequence has provided micro-contextualised insights into the use of the site by hominins and other animals. These new data largely support previous interpretations based on field observations and other proxy datasets (e.g., faunal and pollen records6,7,8,9,35,59), thus increasing confidence in environmental reconstructions for the cave and surrounding region.
Microscopic evidence for hominin use of the cave is minimal. Decades of excavation have generated significant numbers of stone artefacts6,9 that accumulated over a substantial time interval and represent multiple occupational pulses. Micro-remains of a hominin presence—such as combustion bi-products—are readily mobilised and re-deposited, so the lack of intact features indicating fire-use in the Pleistocene sequence is intriguing. Given the limited spatial area that our micromorphological study encompasses, this outcome could be due to sampling bias (e.g., ref.60). On the other hand, because easily dispersed combustion bi-products are, nonetheless, very rare, the early occupants of the site may not have been prolific pyrotechnologists. Where we do record charcoal, it is usually well preserved, so we can rule out the possibility of complete degradation of this material and its preferential removal from the sedimentary record.
The abundant coprolite record shows that the cave was occupied by animals near-continuously. Cave hyena (Crocuta crocuta speleans)—the dominant carnivore in the Altai during the Pleistocene6,59—was present throughout the period of deposition of the Pleistocene sediments7,9,35. Whereas bones can accumulate at a cave site through the agency of various animals, animal droppings are most likely to be delivered directly to the cave floor. Coprolites, then, can be viewed as authigenic components of the sedimentary fill and we surmise that animals—mainly carnivores—used the sampled area of the cave throughout the time represented by the preserved sediments. Fossils of cave hyena are considerably more frequent in the DCE faunal record than are those of other Pleistocene predators, such as wolf, so hyenas are most likely the main accumulating agents of the faunal remains, given the dominance of their coprolites in the cave sediments.
Coprolite fragments occur in high frequencies in layers that have been affected by frost action (e.g., layers 11.4 and 11.2 in DCE). We cannot rule out the possibility that the sampled areas fall within specific latrine areas used for ‘social defecation’61,62, perhaps exacerbated by colder temperatures driving the animals into the further recesses of the cave. At Zhoukoudian Cave in northeastern China60, rich coprolite concentrations and signs of trampling were recognised from sampling localities close to the walls of the cave, confirming the attraction of these animals to marginal zones. Profiles sampled close to the walls of caves may, therefore, fall in areas favoured for animal latrines, underscoring the importance of sampling at multiple locations throughout a site.
An important outcome of our study is the identification of microstratigraphic features consistent with freezing conditions. At present, even on a sunny summer’s day, the cave interior is cold, especially so in DCE, due to the high thermal mass of the surrounding rock mitigating the warm temperatures experienced outside the cave. The cave sediments are frozen during the winter months, when temperatures can drop to an average of −16 °C in January9, but apparently not to the extent that the sediment fabric is re-arranged by ice lensing. The platy structures developed in layers deposited during late Marine Isotope Stage (MIS) 4 and MIS 3 in DCM, and during MIS 6 and MIS 3 in DCE, suggest colder conditions than those experienced in the current interglacial. We postulate that the platy features observed do not necessarily reflect the coldest conditions at the site during the Pleistocene, but are associated with specific formation environments—cold and humid conditions—that allow ice lensing to occur63.
While we observe incipient platy structures in sediments dated to terminal MIS 3 and early MIS 2 (e.g., layer 9, deposited between about 40 and 20 ka), and thus overlapping with the coldest period of the last glacial cycle, the platiness is best expressed in layers that accumulated during the preceding, comparatively warm MIS 3. There is debate surrounding the synchronicity of glacial expansion in the Altai with global climate proxies64,65, potentially associated with increased moisture availability in central Asia during MIS 3 relative to MIS 2, which was drier. The Denisova Cave microstratigraphic record may, therefore, document both cold and relatively humid conditions66, sufficient to develop platy microstructures in the near-surface, cave floor sediments.
Our study provides spatially resolved information on the depositional and post-depositional environments that have preserved organic materials in Denisova Cave. Throughout the sequence, we record only limited evidence of severe chemical diagenesis in the Pleistocene layers, but these include layers that have also yielded sedimentary aDNA10. Even in these layers, however, the evidence for diagenesis is generally weak, albeit spatially variable, providing opportunities for DNA preservation. Cold conditions may also promote the survival of genetic material and lipids in coprolites, from which aDNA and faecal biomarkers can be extracted directly for species identification and dietary reconstruction67,68.
Conclusions
The deposits in Denisova Cave contain microscopic traces of hominin and animal activities that illuminate the use of the cave over the last three glacial–interglacial cycles (Fig. 3). The micromorphological results show that the cave sediments are predominantly geogenic (naturally occurring), augmented by biogenic (biological) additions (e.g., coprolites, guano and digested bone) and anthropogenic inputs (e.g., charcoal, stone artefacts and associated debitage).
Relationships between the various lines of evidence (e.g., micro-charcoal, bioturbation, coprolites and diagenesis), examined at a finely resolved spatial scale, reveal that hominin activities in the microstratigraphic record are few. On the other hand, coprolitic evidence for cave-dwelling carnivores is ubiquitous and suggests that the site often served as a den for hyenas and, to a lesser extent, for wolves.
The cave was visited sporadically by hominins, who appeared not to have been prolific users of fire, at least in the Middle Palaeolithic deposits that constitute the majority of the Pleistocene sequence. The low frequency of hominin occupation has implications for determining the potential agency by which the few Denisovan and Neanderthal fossils were introduced to the site and their post-depositional stratigraphic integrity9,12.
The environmental conditions that best preserve organic molecules, such as DNA and lipids, and whether these materials can be recovered from specific components of the microstratigraphy (e.g., coprolite fragments), also warrant further investigation. Ongoing work at Denisova Cave aims to more fully integrate micromorphology and sedimentary aDNA analyses to develop a predictive tool for organic material preservation in the deposits at this unique hominin locality.