|
|
Post by Admin on Nov 11, 2020 23:24:52 GMT
 This summer brought the richest harvest of anthropological discoveries to archeologists working at the world-famous Denisova Cave in the south of Siberia. The team of Novosibirsk Institute of Archeology and Ethnography worked in the lowest - the oldest - layers of culture-containing soil in the southern gallery of the cave, dating to 300,000 years ago.  Two teeth - a milk and a molar - were both found within that layer, with the milk tooth discovered at its bottom which would date it to approximately 250,000 years, and the molar found at the top of the layer, with the approximate dating from 170,000 to 190,000 years. Both teeth belonged to Denisovans - an extinct group of ancient human, that lived across Asia during Lower and Middle Paleolithic times, said head of the Denisova Cave expedition Mikhail Shunkov.  It was inside the Denisova Cave in 2008 that Siberian scientists discovered a tiny finger bone fragment of an 'X woman', a juvenile female believed to have lived around 41,000 years ago. Analysis showed she was genetically distinct from thick-browed Neanderthals and modern humans. The recent addition to the human family tree was christened Denisovan.  The outstanding finds have definitively confirmed the earlier theory that the Denisovans migrated from the Middle East about 300,000 years old, Mikhail Shunkov said. Two fragments of the bones were also unearthed this summer in layers 14 and 13, which means they are aged 130 and 120,000 years. Since remains of the Neanderthal people were found in these layers earlier, further analyses is needed to determine is the bones belonged to them or to the Denisovans.  The Denisova Cave lies at the border of the Altai region and the Altai Republic in the south of Western Siberia. Locals call it Ayu Tash, which means the Bear Rock. The world famous cave first caught attention of Soviet scientists in 1970s, when they found paleo-archeological remains which led to further research. It is relatively small with the floor area of about 270m2.  It has three galleries - the cosy Central Chamber with high, arched ceiling and a hole that lets in natural light, the South Gallery and the East Gallery Some 3,000 generations back this hole must have been a great chimney for our ancestors. The cave is nicely positioned above river Anuy, which must have given all three hominids - the Neanderthals, the Denisovans and the Homo Sapiens - some stunning sunset views over the past 120,000 years.  Now the site now has a permanent research camp, a pride of Novosibirsk Institute of Archaeology and Ethnography. Another sensational discovery brought the world’s attention to the Denisova Cave in August 2018, when the research showed that early humans of different ancestry met, had sex and had children. Analyses of a 90,000 years old bone fragment found in the Denisova cave in 2012 - which likely belonged to a teenage girl- showed that she had a Neanderthal mother and a Denisovan father. This is the first known hybrid of these groups. |
|
|
|
Post by Admin on Nov 12, 2020 19:57:49 GMT
At least three hominins (Denisovans, Neanderthals, and early Homo sapiens) and large cave-dwelling carnivores (hyena, wolves, and even bears) used Denisova Cave over a span of 300,000 years, according to researchers who analyzed charcoal from ancient fires, coprolites (fossil animal droppings) and bone fragments from the famous cave site. Their results appear in the journal Scientific Reports. Situated in the foothills of the Altai Mountains in southern Siberia, Denisova Cave occupies a prominent place in world prehistory due to the hominin fossils and ancient DNA recovered from the site. 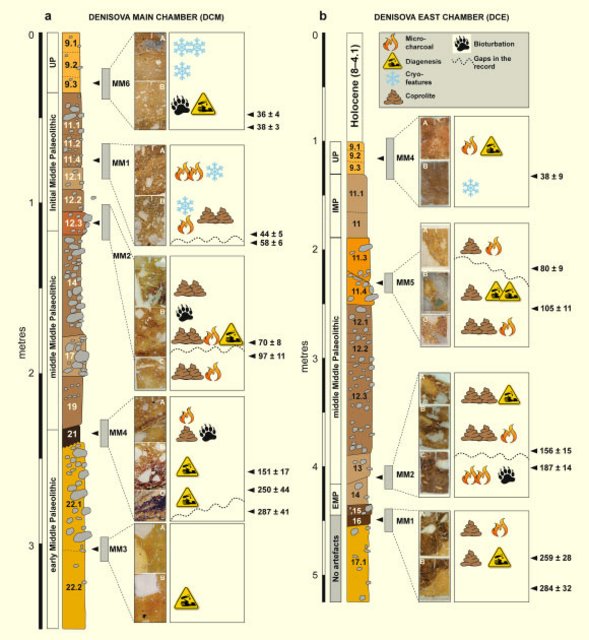 These finds revealed two archaic human populations: Denisovans, identified through genome sequencing of fossils; and Neanderthals, who are at the eastern limits of their known range at this site. “These hominin groups and large carnivores such as hyenas and wolves left a wealth of microscopic traces that illuminate the use of the cave over the last three glacial-interglacial cycles,” said study lead author Dr. Mike Morley, a researcher at Flinders University and the University of Wollongong. “Our results complement previous work by some of our colleagues at the site that identified ancient DNA in the same dirt, belonging to Neanderthals and a previously unknown human group, Denisovans, as well as a wide range of other animals,” he said. “But it now seems that it was the animals that mostly ruled the cave space back then.” From their ‘micromorphology’ examination of the dirt found in the cave, the scientists discovered clues about the use of the cave, including fire-use by ancient humans and the presence of other animals. “This study is very significant because it shows how much can be achieved by sifting through sedimentary material using advanced microscopy and other archaeological science methods to find critical new evidence about human and non-human life on Earth,” said University of Wollongong’s Richard (Bert) Roberts, co-author of the study. “Using microscopic analyses, our latest study shows sporadic hominin visits, illustrated by traces of the use of fire such as miniscule fragments, but with continuous use of the site by cave-dwelling carnivores such as hyenas and wolves.” “Coprolites indicate the persistent presence of non-human cave dwellers, which are very unlikely to have co-habited with humans using the cave for shelter.” “This implies that ancient groups probably came and went for short-lived episodes, and at all other times the cave was occupied by these large predators.” _____ Mike W. Morley et al. 2019. Hominin and animal activities in the microstratigraphic record from Denisova Cave (Altai Mountains). Scientific Reports 9, article number: 13785; doi: 10.1038/s41598-019-49930-3 |
|
|
|
Post by Admin on Nov 13, 2020 19:48:51 GMT
Hominin and animal activities in the microstratigraphic record from Denisova Cave (Altai Mountains, Russia) Mike W. Morley, Paul Goldberg, Vladimir A. Uliyanov, Maxim B. Kozlikin, Michael V. Shunkov, Anatoly P. Derevianko, Zenobia Jacobs & Richard G. Roberts Scientific Reports volume 9, Article number: 13785 (2019) Abstract Denisova Cave in southern Siberia uniquely contains evidence of occupation by a recently discovered group of archaic hominins, the Denisovans, starting from the middle of the Middle Pleistocene. Artefacts, ancient DNA and a range of animal and plant remains have been recovered from the sedimentary deposits, along with a few fragmentary fossils of Denisovans, Neanderthals and a first-generation Neanderthal–Denisovan offspring. The deposits also contain microscopic traces of hominin and animal activities that can provide insights into the use of the cave over the last 300,000 years. Here we report the results of a micromorphological study of intact sediment blocks collected from the Pleistocene deposits in the Main and East Chambers of Denisova Cave. The presence of charcoal attests to the use of fire by hominins, but other evidence of their activities preserved in the microstratigraphic record are few. The ubiquitous occurrence of coprolites, which we attribute primarily to hyenas, indicates that the site was visited for much of its depositional history by cave-dwelling carnivores. Microscopic traces of post-depositional diagenesis, bioturbation and incipient cryoturbation are observed in only a few regions of the deposit examined here. Micromorphology can help identify areas of sedimentary deposit that are most conducive to ancient DNA preservation and could be usefully integrated with DNA analyses of sediments at archaeological sites to illuminate features of their human and environmental history that are invisible to the naked eye. Introduction Situated in the foothills of the Altai Mountains in southern Siberia, Denisova Cave (51°23′51″N, 84°40′36″E; Fig. 1) occupies a prominent place in world prehistory due to the hominin fossils and ancient DNA (aDNA) recovered from the site and the sediments preserved within. These finds have revealed two Pleistocene archaic human populations: a hitherto unknown hominin group, the Denisovans, identified chiefly through genome sequencing of fossils; and Neanderthals, who are at the eastern limits of their known range at this site1,2,3. The hominin story that has emerged from this cave since publication of the Denisovan genome has stimulated considerable scientific debate and media attention. Much of this deliberation has centred on the role of the Denisovans in the recent evolution of our own species and, more broadly, to the demographic composition of Late Pleistocene Eurasia4,5. Figure 1  Archaeological research has been ongoing at Denisova Cave for four decades, during which a great deal of data has been generated regarding the nature and timing of occupational pulses, and the environmental and ecological context of site use6,7,8,9. The cave is situated above the right bank of the Anui River at a point where the valley narrows. The present-day opening of the cave is ~30 m above the modern-day river level, penetrating into the Silurian limestone bedrock along a series of interconnected karstic chambers and tubes that are currently filled with both natural and cultural sediments. Recent excavations of the three principal chambers (Main, East and South Chambers) continue to demonstrate the complexity and spatial heterogeneity of the deposits, revealing a stratigraphic sequence created by geological (e.g., physical and chemical weathering, subsidence and deformation), biological (e.g., animal activity) and anthropogenic (e.g., stone artefact manufacture) processes9. Although the cave remains in the spotlight because of the hominin fossils and aDNA found at the site10,11,12, these items account for only a small fraction of the total material recovered. Much additional information about their immediate and regional context can be gleaned from the study of the deposits that encase them and the cultural remains. A new program of geoarchaeological investigation was initiated in 2014, aimed at documenting the microstratigraphic aspects of the depositional and post-depositional environments represented in the deposits. This work employs micromorphology and contextually-specific elemental mapping of the sediments to seek trace-evidence of processes that formed the site and obtain insights into the behaviours and activities of the site’s occupants—both hominins and other animals—and contextualise at high resolution the artefacts, fossils and genetic material. In particular, our purpose is to: (i) investigate micro-scale evidence for occupation of the cave by hominins and other animals that is not visible at the field scale (e.g., microscopic traces of fire-use, trampling, denning and middening); (ii) further refine our understanding of the genesis of the deposits, including the naturally occurring (geogenic) inputs and post-depositional modifications, and potential correlation to micro-environments in the cave; and (iii) provide an enhanced sedimentary context for the artefacts and for the fossils and sediments from which aDNA has been extracted1,2,3,9,10,11,12. The latter is of special interest given the high degree of aDNA preservation at Denisova Cave. Here, we report the micromorphological results for nine undisturbed sediment blocks collected in 2014 from the Main and East Chambers (DCM and DCE, respectively). Micromorphological analyses retain micro-scale physical relationships13, whereas other analytical techniques commonly employed in archaeological investigation (e.g., zooarchaeological, palynological and stone tool analyses) require the removal of material from its original context (i.e., from the sedimentary matrix) before analysis, resulting in the loss of critical contextual information and associations with complementary datasets. The deposits considered here partly fill DCM and DCE, solution cavities that were formed early in the cave’s history under phreatic and hydrothermal conditions6, most likely at a time well before exposure of the cave to the sub-aerial environment during and after downcutting of the Anui River14. The sediments in DCM and DCE have been described previously from field observations and sedimentological analyses6,7,9,14,15, with the earliest stone artefacts recovered from sediments deposited ~300 thousand years ago (ka)9. The sediments exposed in DCM (Fig. 2a,b) consist largely of fine-grained silts and clays containing angular limestone clasts, with occasional beds dominated by coarser limestone gravel. The lithological layers commonly exhibit abrupt bounding surfaces that are horizontal towards the upper part of the profile, whereas in the lower parts of the sequence they tend to be contorted and convoluted. The lowest unit in DCM (layer 22) has been deformed and subsequently planed by low-energy humid colluviation as seeping water entrained fine-grained sediments producing localised gullying. This process was initiated by slumping, subsidence and plastic deformation, forming two concave depressions, and it is this topographic template that has influenced much of the earlier phases of subsequent sediment deposition. Figure 2b shows the locations of the five micromorphological samples analysed from DCM. |
|
|
|
Post by Admin on Nov 14, 2020 4:16:59 GMT
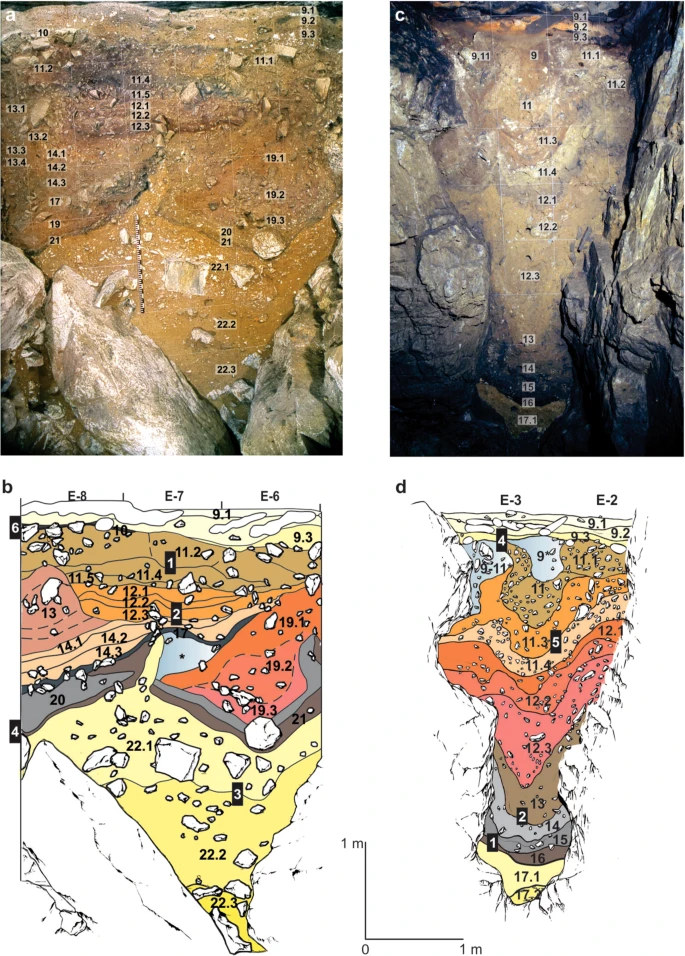 Figure 2 Stratigraphic sequences exposed in the Main (DCM) and East (DCE) Chambers of Denisova Cave. (a) Southeast profile in DCM after excavations in 1984 and (b), Locations of sediment block samples collected in 2014. The concavities on either side of deformed lighter yellow layer 22 (which contains the lowermost artefacts) are infilled with sediments (layers 21–14) that are, in turn, overlain by sub-horizontal layers 13 to 9. (c) Southeast profile in DCE after excavations in 2013 and (d) Locations of sediment block samples collected in 2014. The Pleistocene sequence of hominin occupation spans layers 15 to 9. Layers 17–11 sag towards the centre, above the choke point, and are overlain by sub-horizontal layer 9. Black boxes in panels (b,d) indicate the positions of the micromorphology samples, and the corresponding sample numbers are inset in white; each sample produced 2–3 thin sections, labelled alphabetically from top to bottom. DCM micromorphology samples 4 and 6 were collected from the adjacent east profile. Full size image The sediments preserved in DCE (Fig. 2c,d) are more fine-grained than those in DCM, with many layers significantly enriched with clay. DCE comprises a lower, remnant phreatic tube filled with archaeologically sterile silts and clays, and an upper chamber filled with sediments that contain evidence of both hominins and other animals; the two parts are connected by a narrow slot or choke point. The interfaces between adjacent lithological layers in DCE are convolute, with the sediments centred above the phreatic tube showing downward slumping towards this feature. Figure 2d shows the locations of the four micromorphological samples analysed from DCE. Results Hominin activity in the Denisova Cave microstratigraphic record Hominin fossils and aDNA have been recovered from the sediments preserved at Denisova Cave1,2,3,9,10,11,12, as well as significant numbers of stone artefacts and faunal remains, specimens of which show signs of human modification6,7,8,9,12,16,17. Optical ages9 indicate slow rates of net sedimentation, with periods of non-deposition or erosion, resulting in the accumulation of up to ~4.5 m of Pleistocene deposit in DCM and DCE since ~300 ka (excluding unconformities). Unequivocal signs of hominin activity in the sediments at the field scale are limited, including evidence for fire-use18,19,20,21,22,23 in the Middle Palaeolithic deposits that form the majority of the sequence. We sought microscopic evidence of hominin activity in the sediments, where diagnostic features invisible to the naked eye might be recognised. While we do not identify intact combustion features—common elements of Palaeolithic cave sites—we do observe disassembled combustion bi-products, including micro-charcoal, charcoal fragments and occasional localised ashes. Micromorphological descriptions of all samples examined in this study are provided in Supplementary Information (Table S1), together with a selection of photomicrographs of the thin sections (Fig. S1) and flatbed scans of the sediment blocks (Fig. S2). In DCM (Fig. 3a), we observe micro-charcoal in the basal region within layer 20 (which contains early Middle Palaeolithic artefacts and finished accumulating 170 ± 19 ka), at the interface with overlying layer 19, and as a distinct band within layer 19 (which contains middle Middle Palaeolithic artefacts and started accumulating 151 ± 17 ka). (Archaeological phases and ages, with uncertainties expressed at the 95.4% confidence interval, are from ref.9 (Fig. 3 and Extended Data Table 1) and shown here in Fig. 3 and Table S1.) This micro-charcoal is most likely a taphonomic concentration of combustion bi-products, given the undulating topography of this part of the cave produced by deformation, the truncation of layer 22 by low-energy colluviation, and the localised concentrations of fine charcoal. Layer 19 has produced a total of 1,925 stone artefacts9, so clearly hominins were present at this time. Given the slow sedimentation rate, the artefact assemblage may perhaps represent the product of periodic visitations over many millennia. Site conditions would not have been attractive for hominin occupation during the deposition of these lower layers, owing to the irregular surfaces and occasionally humid conditions in the cave. Higher up the DCM sequence, layers 11.4 and 11.2 also contain charcoal, with larger fragments recorded in layer 11.4 and much finer charcoal powder in layer 11.2. Both layers are associated with the Initial Upper Palaeolithic9. |
|
|
|
Post by Admin on Nov 14, 2020 22:09:03 GMT
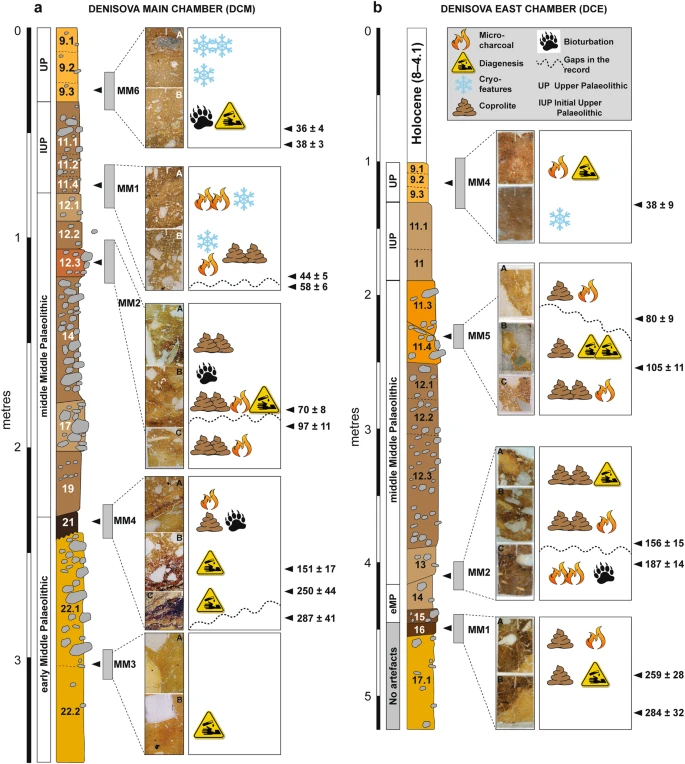 Figure 3 Summary stratigraphic logs of the sequences exposed in (a) DCM and (b) DCE, showing the locations of the micromorphological samples and key microstratigraphic features. To the right of each log, optical ages (in ka, with uncertainties at 95.4% probability) are shown for the major boundaries between lithological units in the thin sections, together with the associated archaeological phases (from ref.9). In DCE (Fig. 3b), we record trace quantities of fine charcoal fragments and flecks closely associated with crushed charcoal and bone fragments in layer 16, and more commonly in layer 15—the earliest layer containing artefacts (early Middle Palaeolithic)9 and also Denisovan DNA10. This indicates fire use and trampling occurring tentatively from 259 ± 28 ka and certainly from 203 ± 14 ka, but we cannot unequivocally and directly link the manufacturers of the stone tools with fire-use because these fine combustion products are highly mobile. In layer 14, which yielded Neanderthal DNA and early Middle Palaeolithic artefacts and was deposited between 193 ± 12 and 187 ± 14 ka, there is a marked increase in micro-charcoal that imparts a dark colouration, and also a small piece (3–4 mm) of angular chert debitage in our sample. We identify large charcoal fragments (>4–5 mm) in layers 11.4 and 11.3 in DCE, which were deposited between 120 ± 11 and 70 ± 8 ka and contain middle Middle Palaeolithic artefacts. A Neanderthal toe phalanx (Denisova 5) was recovered from layer 11.4, but we cannot confidently associate this and other similarly small and isolated hominin fossils with elements of the sedimentary matrix, given the possibility of displacement9,12. In thin section, we observe sediment movement in the form of micro-faulting and slippage features in these layers, presumably associated with the aforementioned post-depositional subsidence. However, such deformation processes do not necessarily promote the translocation or mixing of fine material across lithological boundaries. Our sample location was close to the rear of DCE, where the chamber tapers to a narrow slot (~1 m wide), a locale unlikely to have been conducive to human occupation—especially the lighting of a fire—given the confined space. Evidence of sediment compaction does suggest compressive forces, however, so animals that are represented in the faunal record may have been present in this restricted space. Ashes are present in very low quantities in the Denisova Cave microstratigraphy. Nevertheless, we cannot rule out ash dissolution as the biasing factor, given the mobility of calcite and decalcification recorded locally in some layers. Could fire have been used at Denisova Cave more extensively by hominins, but with the associated evidence subsequently removed from the stratigraphic record? Bearing in mind the rate of cave sedimentation, erosional (chronological) gaps and the evidence for bioturbation in some parts of the sequence—mostly parts of the upper layers of DCE—reworking and redistribution of combustion bi-products may have occurred, although it is unlikely that all micro-traces would have been completely removed. The reworking of previously in situ fire residues is supported by the absence of structured combustion features that would signify the presence of intact hearths. Furthermore, stone tools do not exhibit signs of thermal alteration24, which might be expected should fire-use have been common—or even present—in these confined spaces, and other indicators of fire, such as thermally altered clays, were not evident in our samples. Although fire may not have been used extensively within the sampled areas of the cave, the lack of an obvious pyrotechnology need not preclude the use of a site by hominins, even during glacial periods25. Elsewhere in the Altai, the site of Kara-Bom contains well-preserved hearths in the Initial Upper Palaeolithic deposits26, but no clear evidence of fire-use has been found in the region beyond about 50–40 ka27. Overall, the microstratigraphic record for Denisova Cave indicates that human activity was intermittent over the past three glacial–interglacial cycles represented by the Pleistocene sedimentary infill (>300 ka to ~20 ka). The stone artefact assemblages indicate long-term hominin occupation of the site during both warm climates and cold conditions, when the foothills of the Altai Mountains likely served as a refugium28. Other animal users of Denisova Cave: the fossil coprolites Coprolites are common biogenic components of the cave sediments, often present in dense concentrations, suggesting that animals visited the site for much of its depositional history. The coprolites can be grouped into a number of recurring types throughout the sequence, presumably reflecting the use of the site by a variety of animals, and potentially associated with a range of preservation states. We recognise four main coprolite types (CT-1 to CT-4), described in Table 1 and shown in Fig. 4. Although we cannot confidently attribute all of these droppings to a specific animal, we assign CT-1 to Crocuta crocuta spelaens (cave hyena), based on consistency with published results describing the morphology and optical properties of this material in thin section29,30,31; this supports the faunal evidence of regular use of the cave by these animals6,7,8,9. We tentatively attribute CT-2 to wolf (Canis lupus), based on the similarity between these coprolite fragments and dog coprolites recorded at Vanguard Cave, Gibraltar32, as well as other published data31. The coprolite fragments (CT-3) in our thin section of layer 12.2 in DCM are consistently larger and darker than CT-1 and CT-2. This layer contains very high proportions of these coprolites, and the chaotic arrangement of the coarse limestone gravel, with long axes in a vertical to sub-vertical alignment, suggests disturbance of these sediments, possibly by a large animal such as a cave bear. We cannot assign CT-3 or CT-4 to a specific species. Figure 4 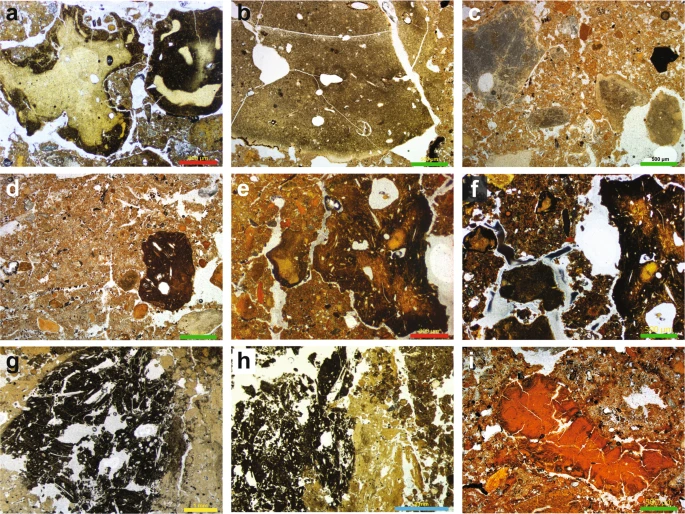 Examples of coprolites identified in the Denisova Cave microstratigraphic record (see Table 1 for coprolite descriptions). (a–c) Type CT-1 originating from cave hyena occurs through much of the sampled sequence; (d–f) Type CT-2 has a much darker matrix, possibly related to wolf; (g,h) Type CT-3 is highly weathered; (i) Type CT-4 has a distinctive red matrix. Types CT-3 and CT-4 cannot be linked to specific animals. Scale bars: red, 800 µm; green, 500 µm; yellow, 1 mm; blue, 2 mm. Coprolite fragments commonly occur in layers that also contain stone artefacts. Given that hominins and hyenas will not cohabitate7,33, this indicates that specific occupation events will be extremely challenging—if not impossible—to tease out at Denisova Cave, with the slow sedimentation rates effectively precluding the identification of alternating hominin–carnivore occupations, should they exist. The lack of defined stratigraphy within layers (e.g., buried surfaces) may be due, at least in part, to carnivore denning or other animal burrowing activities in parts of the deposit. At Bois Roche in France, for example, stone tools accumulated by local movement (e.g., by gravity) in areas that functioned primarily as carnivore dens34. The DCE faunal record includes the remains of a number of large cave-dwelling mammals recovered in relatively high numbers, including hyena, wolf, red fox and, to a lesser extent, bear9,35. As these animals are unlikely to have cohabitated, either with each other or with hominins, the co-occurrence of their remains likely reflects the scale of resolution (time averaging) of the sedimentological, chronological and hominin occupation records at Denisova Cave. The presence of coprolites in layers from which hominin remains and aDNA have been recovered implies that large carnivores might be an accumulating agent for these materials, particularly in areas of the site where evidence for hominin activity is scarce (e.g., the farthest recesses of DCE). Specific areas of the site might have been designated as waste dumps for lithic debitage and food detritus, for example, which in turn attracted scavengers such as the cave hyena when hominins were absent from the site. Interestingly, we record in thin section only a few examples of bone fragments that exhibit characteristic etching related to digestion in the gut of a carnivore, although etched bones are common in the faunal record35. In the field, rodent burrows (krotovinas36) are clearly visible in the Holocene deposits of DCE, and in fewer numbers in DCM. Disturbance of the sediments by bioturbation is also evident in thin section. Parts of layer 12 and much of layer 13 in DCM display a chaotic arrangement of limestone clasts within finer material, consistent with disturbance by large animals such as bears, wolves or hyenas. This accords with field observations of layer 13 being a hyena lair9. Thin sections of layers 9.2 and 9.3 in DCM and layers 9.1, 11.3 and 11.4 in DCE display abundant, loosely arranged aggregates and irregular vughs typical of bioturbation37. These small, mm-size features are typical of smaller soil fauna, such as worms, spring-tails (Collemboles) and isopods. We note that these fine crumb structures occur essentially in the uppermost Pleistocene strata in DCE (i.e., layer 9), which accumulated after 38 ± 9 ka and may represent milder conditions that enabled these fauna to flourish. |
|

