|
|
Post by Admin on Jun 23, 2021 20:16:42 GMT
 "Just collecting hundreds of samples from all three chambers in the cave, and documenting their precise locations, took us more than a week, but we obtained a comprehensive set of samples spanning more than 300,000 years of Siberian history," Professor Jacobs said. The new study builds on the detailed timeline obtained from optical dating of the Denisova Cave sediments at UOW, published in Nature in 2019. "The chronology generated previously for the cave sediments allowed us to choose the best places to collect the DNA samples and make the most of the extraordinary insights from sediment DNA," Professor Jacobs said. At the Max Planck Institute for Evolutionary Anthropology in Leipzig, Ph.D. student Elena Zavala, the study's lead author, extracted and sequenced small traces of ancient human and animal mitochondrial DNA from the huge collection of samples. "These efforts paid off and we detected the DNA of Denisovans, Neanderthals and ancient modern humans in 24 percent of the samples," she said. When matching the DNA profiles with the ages of the layers, the researchers found the first humans to visit the site were Denisovans about 250,000 years ago, followed by Neanderthals about 200,000 years ago. Only Neanderthal DNA was found in sediments deposited between 130,000 and 80,000 years ago. Denisovans who came back after this time carried a different mitochondrial DNA to Denisovans who were there earlier, suggesting a different population had arrived in the region.  Modern human DNA first appears in the Initial Upper Palaeolithic layers, which contained pendants and other ornaments made from animal bones and teeth, mammoth ivory, ostrich eggshell, marble and gemstones. "This provides not only the first evidence of modern humans at the site, but also suggests that they may have brought new technology into the region with them," Ms Zavala said. The scientists also found animal DNA in nearly all samples, and identified two time periods when changes occurred in both animal and human populations. The first, around 190,000 years ago, coincided with a shift from relatively warm to relatively cold conditions, when hyaena and bear populations changed and Neanderthals first appeared in the cave. The second major change occurred between 130,000 and 100,000 years ago, along with a shift in climate from relatively cold to relatively warm conditions. During this period, animal populations changed again, Denisovans vanished, and Neanderthals were left as the only human occupants of the cave. "The coincidence of these population turnovers with climatic transitions between interglacial and glacial periods suggests that environmental factors played a key role in shaping the human and faunal history of this region," Professor Roberts said. Professor Matthias Meyer, leader of the Advanced DNA Sequencing Techniques group at the Max Planck Institute for Evolutionary Anthropology and senior author of the new study, said: "Being able to generate such dense genetic data from an archaeological site is like a dream come true. There is so much information hidden in sediments—it will keep us and many other geneticists busy for a lifetime." Explore further New studies reveal deep history of archaic humans in southern Siberia More information: Pleistocene sediment DNA reveals hominin and faunal turnovers at Denisova Cave, Nature. doi.org/10.1038/s41586-021-03675-0 |
|
|
|
Post by Admin on Jun 24, 2021 3:01:21 GMT
Pleistocene sediment DNA reveals hominin and faunal turnovers at Denisova Cave Elena I. Zavala, Zenobia Jacobs, Benjamin Vernot, Michael V. Shunkov, Maxim B. Kozlikin, Anatoly P. Derevianko, Elena Essel, Cesare de Fillipo, Sarah Nagel, Julia Richter, Frédéric Romagné, Anna Schmidt, Bo Li, Kieran O’Gorman, Viviane Slon, Janet Kelso, Svante Pääbo, Richard G. Roberts & Matthias Meyer Abstract Denisova Cave in southern Siberia is the type locality of the Denisovans, an archaic hominin group who were related to Neanderthals1,2,3,4. The dozen hominin remains recovered from the deposits also include Neanderthals5,6 and the child of a Neanderthal and a Denisovan7, which suggests that Denisova Cave was a contact zone between these archaic hominins. However, uncertainties persist about the order in which these groups appeared at the site, the timing and environmental context of hominin occupation, and the association of particular hominin groups with archaeological assemblages5,8,9,10,11. Here we report the analysis of DNA from 728 sediment samples that were collected in a grid-like manner from layers dating to the Pleistocene epoch. We retrieved ancient faunal and hominin mitochondrial (mt)DNA from 685 and 175 samples, respectively. The earliest evidence for hominin mtDNA is of Denisovans, and is associated with early Middle Palaeolithic stone tools that were deposited approximately 250,000 to 170,000 years ago; Neanderthal mtDNA first appears towards the end of this period. We detect a turnover in the mtDNA of Denisovans that coincides with changes in the composition of faunal mtDNA, and evidence that Denisovans and Neanderthals occupied the site repeatedly—possibly until, or after, the onset of the Initial Upper Palaeolithic at least 45,000 years ago, when modern human mtDNA is first recorded in the sediments. Main Denisova Cave consists of three chambers (designated Main, East and South Chambers) that contain deposits with stratigraphic sequences extending from the Middle Pleistocene to the Holocene epoch. The Pleistocene deposits have chronologies that have been constructed from the radiocarbon dating of bone, tooth and charcoal5 (to around 50 thousand years ago (ka)) and optical dating of sediments8 (to more than 300 ka). Optical ages for Main and East Chambers (Fig. 1a–c) can be aligned on a common time scale (Extended Data Fig. 1) but excavations are ongoing in South Chamber, where layers are only tentatively recognized. Mitochondrial DNA and nuclear DNA have been recovered from eight hominin fossils, enabling four to be assigned to Denisovans (Denisova 2, Denisova 3, Denisova 4 and Denisova 8)1,2,3,4, three to Neanderthals (Denisova 5, Denisova 9 and Denisova 15)5,6,12, and one to the child of a Neanderthal and a Denisovan (Denisova 11)7. However, there are too few fossils to enable the detailed reconstruction of the timing and sequence of hominin occupation, and the association of the early Middle Palaeolithic, middle Middle Palaeolithic and Initial Upper Palaeolithic assemblages identified at the site with specific hominin groups. Moreover, two Denisovan fossils (Denisova 3 and Denisova 4)—but no modern human remains—have been recovered from the Initial Upper Palaeolithic layers, so it is debated whether archaic hominins or modern humans created the associated ornaments and bone tools9,10,11. Fig. 1: Stratigraphic sequences in Denisova Cave, showing locations of sediment samples collected for mtDNA analysis and results obtained for ancient hominins. 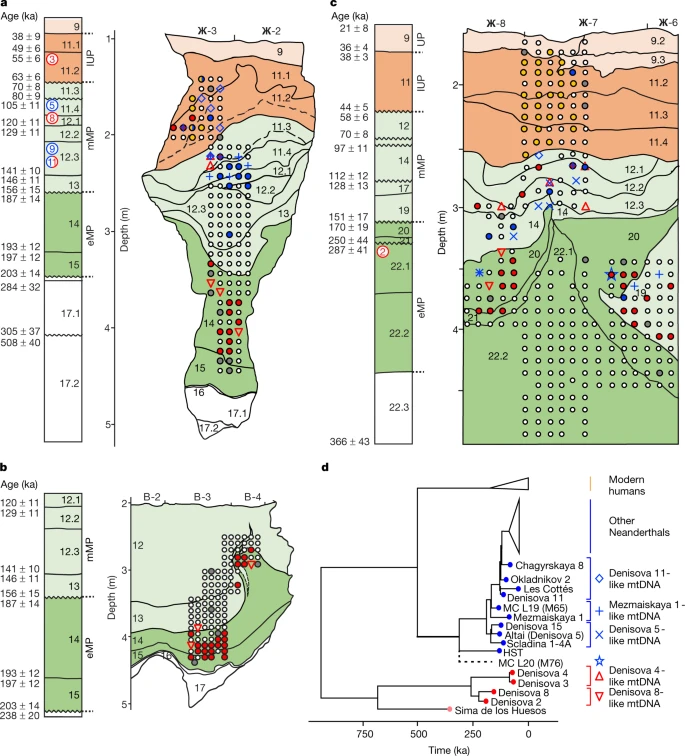 a, East Chamber, southeast profile. b, East Chamber, northwest profile. c, Main Chamber, southeast profile. d, Phylogenetic tree of mtDNA genomes used as references to distinguish specific hominin lineages, and estimated placement of the Neanderthal mtDNA lineage identified in sample M76 from Main Chamber layer 20. Filled circles in a–c indicate the locations of individual sediment samples, and colours correspond to the hominin mtDNA detected: red (Denisovan), blue (Neanderthal), yellow (ancient modern human), grey (unidentified ancient hominin) and white (no ancient hominins detected). Other symbols denote samples for which mtDNA could be assigned to one of the specific hominin lineages in d (Denisovan, red open triangles; Neanderthal, blue open diamonds, crosses and star). Composite stratigraphic sections (modified from a previous publication8) to the left of each profile show modelled start and end ages (in ka) for sedimentary layers (uncertainties at 95.4% probability). Hominin specimen numbers are shown circled in the layer from which each fossil was recovered: Denisovan (red), Neanderthal (blue) and Neanderthal–Denisovan offspring (both colours)5. Dashed lines in profiles in a and b indicate areas in which layer assignment is uncertain8. Background shading denotes the associated archaeological assemblage: early Middle Palaeolithic (eMP) (dark green), middle Middle Palaeolithic (mMP) (light green), Initial Upper Palaeolithic (IUP) (dark orange) and Upper Palaeolithic (UP) (light orange). A pilot study of DNA preservation in sediments from Denisova Cave identified ancient hominin mtDNA in 12 out of 52 samples13, which suggested a path to reconstructing the occupational history of the site at higher resolution than is feasible from the scarce hominin fossil record. Here we report the analysis of 728 sediment samples, collected in a 10–15-cm grid-like pattern from the exposed Pleistocene deposits in all three chambers (Extended Data Figs. 2, 3a, b, Supplementary Information sections 1, 2). Using automated laboratory protocols, DNA was extracted from each sample, converted to single-stranded libraries and enriched for mammalian and hominin mtDNA13,14, which we identified to the biological-family level using an established analysis pipeline13. |
|
|
|
Post by Admin on Jun 24, 2021 21:24:41 GMT
Ancient hominin mtDNA We detected ancient hominin mtDNA in 175 samples (24%), covering nearly all layers in all three chambers (Fig. 1a–c, Extended Data Figs. 3a, b, 5b). Four samples showed evidence for the presence of predominantly one haplotype and yielded sufficient mtDNA fragments to reconstruct mtDNA consensus sequences that are more than 80% complete (Supplementary Information sections 5, 6). Three of the sequences (samples E202 and E213 from East Chamber (layers 11.4 and 11.4/12.1) and sample M65 from Main Chamber (layer 19)) group with Neanderthals in phylogenetic trees built with previously published hominin mtDNAs (Supplementary Information section 7), specifically with Denisova 5, Denisova 15, Mezmaiskaya 1 and Scladina I-4A (Fig. 1d, Extended Data Fig. 6a). The fourth sequence (sample M71 from Main Chamber (layer 20)) is of the Denisovan type and falls basal to Denisova 2 and Denisova 8, albeit with low bootstrap support (Extended Data Fig. 6b). The most complete mtDNA sequence (over 99% of the genome reconstructed) for a Neanderthal from Main Chamber (M65) has a genetic age estimate of 140 ka (95.4% highest posterior density interval of 181–98 ka) (Supplementary Information section 7), consistent with the time of deposition of layer 19 (151 ± 17 to 128 ± 13 ka; here and below, uncertainties on optical ages are given at 95.4% probability)8. For the remaining 171 samples, we assigned mtDNA fragments to specific hominin groups by counting the number of fragments that support lineage-specific states at diagnostic sites that distinguish between modern human, Neanderthal and Denisovan mtDNA genomes. We distinguished three Neanderthal lineages: the Sima de los Huesos lineage (representing Neanderthals who lived approximately 430 ka in Spain and whose mtDNA is most closely related to that of Denisovans)18,19; the Hohlenstein–Stadel (HST) lineage, which falls basal to all other Neanderthal mtDNAs20; and the ‘typical’ Neanderthal mtDNA, known from all other Neanderthals. The presence of ancient modern human mtDNA was evaluated by restricting the analysis to deaminated fragments to mitigate the effect of present-day human DNA contamination. We identified Denisovan and typical Neanderthal mtDNA in 79 and 47 samples, respectively (based on 54–9,093 unique hominin mtDNA fragments), and modern human mtDNA in 35 samples (based on 55–2,200 deaminated fragments) (Fig. 1a–c, Extended Data Fig. 3a, b). We detected DNA from two hominin groups in ten samples, either within a single library or across libraries that were prepared from independent subsamples in some cases (Extended Data Fig. 6c, Supplementary Information section 4). In addition, we identified one sample (M76 from Main Chamber (layer 20)) containing hominin mtDNA fragments that support the branch shared by HST and typical Neanderthal mtDNA, but neither of the branches defining those lineages. This signal cannot be created by mixing mtDNA fragments from Neanderthals, Denisovans and ancient or present-day modern humans. On the basis of simulations with ancestralized Neanderthal mtDNA, the mtDNA in this sample is compatible with the presence of a previously unknown Neanderthal mtDNA lineage that diverged from typical Neanderthal mtDNA between 255 and 230 ka, 20 to 45 thousand years after the split of the HST and typical Neanderthal mtDNA lineages (Supplementary Information section 9). The oldest hominin mtDNA recovered—identified as Denisovan—originates from a sample in Main Chamber layer 21, which began to accumulate 250 ± 44 ka. This provides the earliest genetic evidence for hominin occupation in Denisova Cave; Denisova 2 was found in layer 22.1, but is probably intrusive from an overlying layer and has an estimated age of 194–123 ka5. Among all 223 samples from the early Middle Palaeolithic layers in Main and East Chambers, 50 contained evidence for Denisovan mtDNA and only three (all from layer 20 in Main Chamber) for Neanderthal mtDNA. Two of these (M174 and M235) contain typical Neanderthal mtDNA and are from areas in which small-scale mixing with overlying sediments may have occurred8; the third (M76) is from the middle of the layer, and carries the previously unknown Neanderthal mtDNA lineage. These results point to Denisovans as the first and principal makers of the early Middle Palaeolithic assemblages, which are older than 170 ± 19 ka. Consistent with this interpretation, the detection of Neanderthal mtDNA in a sediment sample from early Middle Palaeolithic layer 14 in East Chamber in the pilot study13 was due to an incorrect assignment, which was later corrected to middle Middle Palaeolithic layer 11.4 in this chamber8 (Supplementary Information section 2). Our results also suggest that Neanderthals first occupied Denisova Cave towards the end of the early Middle Palaeolithic and may therefore have contributed to the production of these assemblages in their later stages. Forty out of 173 samples from the middle Middle Palaeolithic layers in Main and East Chambers (deposited approximately 160–60 ka) yielded Neanderthal and/or Denisovan mtDNA, with both present in six samples (Fig. 1a–c). DNA from both groups also occurs in the deformed Middle Palaeolithic layers in South Chamber (Extended Data Fig. 3b). Notably, sediments deposited between 120 ± 11 and 97 ± 11 ka in Main and East Chambers produced no traces of Denisovan mtDNA, whereas 12 samples contained Neanderthal mtDNA. This suggests that only Neanderthals may have occupied the cave during that period, and possibly for most of Marine Isotope Stage (MIS) 5 (Fig. 2). Fig. 2: Timeline of hominin and faunal mtDNA presence, archaeological phases and environmental records at Denisova Cave. 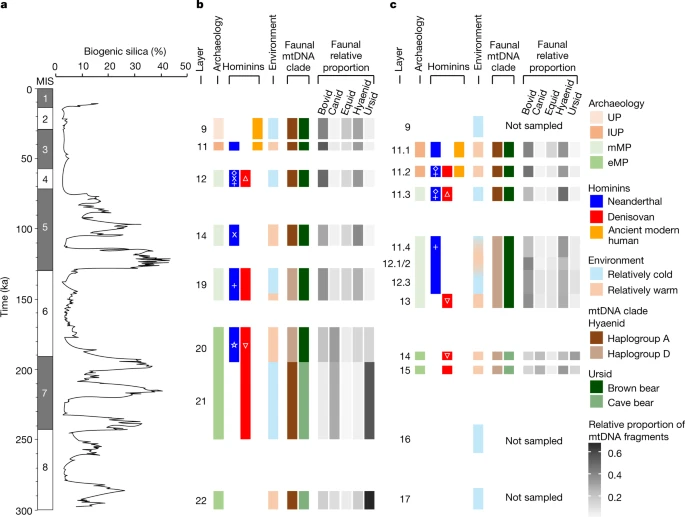 a, Baikal Drilling Project 1996 composite biogenic silica record of diatom productivity in Lake Baikal, a proxy for regional annual temperature28. b, c, Summary data for Main Chamber (b) and East Chamber (c). Start and end ages for layers and corresponding data are based on the common time scale in Extended Data Fig. 1. Time intervals in white represent gaps in the stratigraphic sequence, or were not sampled. Archaeological phases (early Middle Palaeolithic, middle Middle Palaeolithic, Initial Upper Palaeolithic and Upper Palaeolithic) follow the colour scheme in Fig. 1 and Extended Data Fig. 1. Genetic data for hominins are from Fig. 1 (excluding samples M174 and M235 from Main Chamber, which are thought to be out of context), and for dominant hyaenid and ursid populations from Extended Data Fig. 10. Symbols for specific hominin mtDNA lineages (Fig. 1d) are inset in white. Environmental conditions are inferred from pollen records and skeletal remains of vertebrate fauna8. Relative proportions of bovid, canid, equid, hyaenid and ursid mtDNA are from Extended Data Figs. 8, 9. Only ancient modern human mtDNA was detected in the Initial Upper Palaeolithic and Upper Palaeolithic layers in Main Chamber (layers 11.4 and above, deposited 44 ± 5 to 21 ± 8 ka), except for one sample from Initial Upper Palaeolithic layer 11.2 that yielded Neanderthal mtDNA (Fig. 1c). The association between Initial Upper Palaeolithic assemblages and the appearance of modern humans is further supported by the recovery of modern human mtDNA from a sample from Initial Upper Palaeolithic layer 11 in South Chamber, which was deposited after 47 ± 8 ka (Extended Data Fig. 3a). The situation in East Chamber is more complex: Denisovan, Neanderthal and ancient modern human mtDNAs were recovered from Initial Upper Palaeolithic layer 11.2, and Neanderthal and ancient modern human mtDNA from Initial Upper Palaeolithic layer 11.1 (Fig. 1a). Given these results and the recovery of two Denisovan fossils (Denisova 3 and Denisova 4) from layers associated with Initial Upper Palaeolithic assemblages, we cannot discount the possibility that—in addition to modern humans—Denisovans and Neanderthals may have been present during the period of Initial Upper Palaeolithic production9,10,11. For 34 out of 37 samples that yielded 100 or more deaminated hominin mtDNA fragments, we identified similarities with specific Neanderthal and Denisovan mtDNA genomes using a k-mer-based approach21,22 (denoted by symbols other than circles in Fig. 1a–c, Extended Data Fig. 3b, Supplementary Information section 8). This analysis revealed that the early Middle Palaeolithic layers in Main and East Chambers and the earliest middle Middle Palaeolithic layer in East Chamber, which span the period between 250 ± 44 and 146 ± 11 ka, contain Denisova-2- and Denisova-8-like mtDNA fragments. This contrasts with the Denisovan mtDNA recovered from middle Middle Palaeolithic layers deposited after 80 ± 9 ka, which yielded assignments to Denisova-3- and Denisova-4-like sequences, as did a sample from South Chamber. These results suggest a transition in mtDNA sequences sometime in the 146–80-ka interval, possibly reflecting different Denisovan populations. Our results also align well with the modelled ages of Denisova 2, Denisova 3 and Denisova 4, and with the relative age of Denisova 8 inferred from molecular dating5. Sediments deposited between about 130 and 100 ka (and possibly longer, given the subsequent time gap of 20 millennia)—during MIS 5—contain mtDNA and fossil evidence only of Neanderthals, with Denisova-11-like mtDNA sequences only appearing in sediments deposited after 80 ka. |
|
|
|
Post by Admin on Jun 25, 2021 4:41:50 GMT
Ancient faunal mtDNA
All large mammals present in the palaeontological record of Denisova Cave (currently available for Main and East Chambers only8) were also identified in the sediment DNA (Extended Data Fig. 7a, Supplementary Information section 10). In addition, ancient mtDNA from camelids was found in one sample from East Chamber (layer 12), consistent with the Pleistocene presence of Camelus knoblochi in the region23. In contrast to large mammals, small mammals (such as Spalacidae, Leporidae and Sciuridae) are largely absent from the genetic data (Extended Data Fig. 7b), which may be due to their lower biomass or underrepresentation among the capture probes. The sharp boundaries between the faunal mtDNA composition in some of the adjacent layers (for example, between layers 22.1 and 21/20 in Main Chamber) (Extended Data Fig. 8) provide additional evidence that post-depositional leaching of DNA is limited, if it occurs.
Despite the highly fragmentary nature of fossil remains at Denisova Cave8 and the varying quantities of DNA that may be deposited by different species, changes in the relative abundance of mammalian DNA over time are broadly consistent with changes in the skeletal records for some families, such as bovids, hyaenids, ursids and canids (Extended Data Fig. 7c, d). Genetic data also provide the opportunity to study faunal diversity at the species or population level, in cases in which comprehensive reference data are available (as is the case for elephantids, ursids and hyaenids) (Supplementary Information section 11). Elephantid mtDNA was assigned predominantly to woolly mammoth in all layers, whereas the relative abundance of ursid species shifted from predominantly cave bear mtDNA in layers deposited before 187 ± 14 ka to exclusively brown bear mtDNA after 112 ± 12 ka (Extended Data Fig. 10). We also detected the presence of three previously described mtDNA haplogroups of the genus Crocuta (spotted and cave hyaenas)24. Layers deposited before 200 ka and after 120–80 ka contain mostly mtDNAs seen in African spotted hyaenas and European cave hyaenas (haplogroup A), whereas layers of intermediate age contain mtDNA predominantly from east Asian cave hyaenas (haplogroup D) and some from European cave hyaenas (haplogroup B) (Extended Data Fig. 10). The Altai Mountains may therefore have been a contact zone for both hominins and distinct lineages of hyaena and other fauna, as previously suggested by studies of mammalian skeletal remains25.
At least two major turnovers of large mammals are evident from the sediment DNA (Fig. 2). First, marked changes in the relative proportions of mtDNA fragments of bovids, canids, equids, hyaenids and ursids, a turnover in hyaena mtDNA haplogroups and a shift from cave bears to brown bears occurred about 190 ka, contemporaneous with the climatic transition from an interglacial period (MIS 7) to a glacial period (MIS 6). The earliest traces of Neanderthal mtDNA also appear around this time. A second turnover took place between about 130 and 100 (or 80) ka, during and after the climatic transition from MIS 6 to MIS 5: mtDNA proportions of bovids, canids, felids and ursids declined, whereas those of cervids and equids increased, and cave bears and two hyaena haplogroups disappeared (Extended Data Fig. 10). This period is notable also for the absence of Denisovan mtDNA in the cave sediments. These changes suggest that turnovers in hominin and faunal populations may have been linked, and related to ecological factors25.
Discussion
The identification of archaic hominin mtDNA in 175 sediment samples exceeds by an order of magnitude the number of hominin fossils retrieved from the deposits in Denisova Cave and provides a genetic profile of the presence of hominins in nearly all of the Pleistocene layers (Fig. 2). These data are complemented by faunal mtDNA sequences from 685 samples, which provide information about the diversity of other large mammals and changes in their relative abundance. However, we caution that the inferred sequence of hominin and faunal occupation is constrained by several factors: the existence of two major gaps in the stratigraphic record (170–156 and 97–80 ka), the time-averaging inherent in the accumulation of each sediment layer, the post-depositional disturbance of some layers owing to burrowing animals or small-scale mixing8,26, and the precision of the optical ages used to construct the site chronology.
Beyond reconstructing the occupational history of Denisova Cave, our results have wider implications for understanding the human past. First, fragments of Denisovan mtDNA recovered from the middle Middle Palaeolithic layers deposited in Denisova Cave after 80 ka consistently show the highest similarity to the mtDNA of Denisova 3 and Denisova 4, as do mtDNA fragments retrieved from sediments at Baishiya Karst Cave on the northeastern flank of the Tibetan Plateau that are broadly contemporaneous in age (70–45 ka)27. This pattern suggests that this lineage was the most abundant mtDNA type carried by Denisovans after 80 ka. Palaeontological studies25 have suggested that Pleistocene mammals migrated from southeast Asia, along the eastern foothills of the Himalayas, to the northwest Altai. These faunal migrations may have spurred the dispersal of Denisovans into the region in which their remains were first discovered. Second, the presence of Neanderthal mtDNA before 170 ka further constrains the timing of an early event in Neanderthal history—the replacement of the mtDNA lineage found in Neanderthals who lived 430 ka in Spain by mtDNA that introgressed from early ancestors of modern humans18,19—to between 430 and 170 ka. High-resolution profiling of sediment DNA can therefore provide an effective means of filling gaps in our knowledge of human evolutionary history and palaeoecology, independent of the discovery of skeletal remains.
|
|

