|
|
Post by Admin on Mar 5, 2017 20:18:39 GMT
 The origin of the widespread Scythian culture has long been debated in Eurasian archaeology. The northern Black Sea steppe was originally considered the homeland and centre of the Scythians3 until Terenozhkin formulated the hypothesis of a Central Asian origin4. On the other hand, evidence supporting an east Eurasian origin includes the kurgan Arzhan 1 in Tuva5, which is considered the earliest Scythian kurgan5. Dating of additional burial sites situated in east and west Eurasia confirmed eastern kurgans as older than their western counterparts6,7. Additionally, elements of the characteristic ‘Animal Style’ dated to the tenth century BCE1,4 were found in the region of the Yenisei river and modern-day China, supporting the early presence of Scythian culture in the East. Artefacts of the Scythian culture spread over a large territory shortly after its emergence, but the underlying population dynamics that may have driven the cultural diffusion are poorly understood.  Genetic studies on Central Asian populations based on both ancient8 and modern mitochondrial DNA (mtDNA)9,10,11,12 agree that Central Asia has historically been a crossroad for population movements from east to west and vice versa. It has been claimed that gene flow occurred from east to west Eurasia as early as the Palaeolithic13,14 and the Mesolithic15, and from west to east Eurasia during the Bronze Age16. A recent genomic study17 has emphasized the role of eastward migrations of people associated with the Yamnaya and Andronovo culture during the Bronze Age, followed by substantial admixture with East Asians. Most genetic studies on the later Iron Age nomads, however, have been limited by small sample size, restricted to a single cultural group, or based on the analysis of mtDNA alone18,19,20,21,22. In this study, mtDNA data from 96 individuals associated with the Scythian culture in different geographical regions and time periods have been sequenced and analysed; additionally, genomic data from eight of these individuals was obtained and analysed (Supplementary Table 1). From the western part of the Eurasian Steppe, samples discovered in the North Caucasus dating to the initial Scythian period (eighth to sixth century BCE), classical Scythians from the Don-Volga region (third century BCE), and Early Sarmatians from Pokrovka, southwest of the Ural (fifth to second century BCE), were included. From the eastern part of the Eurasian Steppe, we analysed samples from East Kazakhstan dating to the Zevakino-Chilikta phase (ninth to seventh century BCE); from the site Arzhan 2, assigned to the Aldy Bel culture in Tuva (seventh to sixth century BCE); and from the Tagar culture of the Minusinsk Basin (fifth century BCE). The majority of the samples generated by this study or retrieved from the literature date to the fourth to third century BCE and were discovered at archaeological sites situated in the Kazakh, Russian and Mongolian parts of the Altai Mountains. These findings were all assigned to the Pazyryk culture, which is named after the first discoveries by Gryaznov in 1927 and 1929 in the Pazyryk Valley and famous for its rich frost-conserved graves, where human bodies, tapestry and clothing remained well-preserved23,24,25 (Fig. 1).  Figure 1: Reconstruction of a Scythian. While the eastern and western populations investigated here are separated by a distance of 2,000–3,500 km, archaeological evidence indicates that they were strikingly similar regarding their lifestyle and culture. The aims of this study are to investigate the extent to which these groups are genetically related to each other and whether they have a common origin, and to elucidate their demographic history and genetic relationships to modern living populations. We therefore divided the Iron Age steppe nomad data generated by this study and from the literature (n=147, Supplementary Table 1) into seven sample groups (see Fig. 2) based on geographical and chronological criteria, and analysed these ancient individuals together with an extensive sample of modern individuals from 86 populations (n=3,410, Supplementary Fig. 11 and Supplementary Table 19) from all parts of Eurasia. Our analysis included an array of statistical methods as well as a series of population genetic inference approaches, including explicit demographic modelling.  Figure 2: Distribution of the sample sites analysed for this study (yellow) including data from the literature (green). |
|
|
|
Post by Admin on Mar 6, 2017 20:18:25 GMT
Genetic relationship and origin of the Scythian groups The eastern sample group (n=113) can be divided in four cultural subgroups chronologically dispersed over the 1st millennium BCE (Fig. 2). Analysing mtDNA, we found no significant genetic distance between those groups (Supplementary Table 8). We then used approximate Bayesian computation (ABC)28 to test for continuity between the earlier (#4: Zevakino-Chilikta and #5: Aldy Bel; n=26) and the later (#6: Pazyryk; n=71) Scythian period in the East. The Tagar/Tes group (#7) had to be excluded because of their imprecise dating. These analyses revealed that they were most likely derived from one single population that was expanding over the time period considered here, that is, the two samples are unlikely to represent two independent populations that diverged earlier than 108 generations before present (g BP) or ∼2.7 ky BP. This scenario was highly supported in our model selection procedure (Supplementary Table 9, logistic regression, P=0.995, neural networks P=0.568, cf. confidence in model choice in Supplementary Table 10, model parameter posteriors in Supplementary Table 11), whereas two scenarios that assumed that the eastern Scythian sample groups were derived from two previously diverged populations received very little statistical support (cumulative posterior probabilities: logistic regression, P<0.001, neural networks, P=0.001). Since the genetic distance between the combined Scythian groups of the east versus those of the west is relatively low (FST=0.01733; P-value=0.02148±0.0045), we used ABC to further assess if the eastern and western Scythians might share a common origin (Fig. 3). For these analyses we included contemporary samples representative of genetic diversity on the extremes of Eurasia (Supplementary Note 1). According to our model selection algorithm, a multiregional model provided the best fit to the empirically observed diversity patterns (Supplementary Table 12, 0.5% closest simulations, posterior probability P=0.708 for logistic regression, P=0.715 for neural networks method), while a model of western origin also received some support (Supplementary Table 12, 0.5% closest simulations, posterior probability P=0.286 for logistic regression, P=0.267 for neural networks method). Therefore, in contrast to the eastern origin model, a western origin cannot be fully discounted by our analysis. In addition, a pairwise comparison through the computation of Bayes factors reveals a substantial to strong (logistic regression) or weak to substantial (neural network) support for the multiregional origin over the western origin model (see Supplementary Note 1, Supplementary Table 14 and Supplementary Fig. 5b). These results suggest that western and eastern Scythian groups arose independently—perhaps in their respective geographic regions—and thereafter experienced significant population expansions (during the 1st millennium BCE). Importantly, our simulations support a continuous gene flow between the Iron Age Scythian groups, with indications of asymmetrical gene flow from western to eastern groups, rather than the reverse (see Supplementary Tables 13 and 15 for details).  Figure 3: Candidate scenarios for the origin of Scythian populations. Because population movements across Central Asia during the Bronze Age are often archaeologically associated with the spread of the Andronovo culture29, we used ABC to fit a sample of Middle Bronze Age nomadic groups from western Siberia, most of them associated with the Andronovo culture, onto the preferred demographic model for the origin of Scythians. For this purpose—and based on low FST values between these groups—we combined 40 samples related to the Andronovo culture in the west Siberian forest steppe30 and nine samples from the same culture in the Krasnoyarsk region31, all of which were dated to the first half of the 2nd millennium BCE. The results provided very strong support for a linkage between these Middle Bronze Age groups and eastern Scythians (Supplementary Tables 16 and 17). However, these simulations were not able to fully capture the patterns of genetic diversity observed in the Bronze Age populations, suggesting that the true demographic history of the ancestry of Iron Age populations may have been more complex than considered here (see Supplementary Note 1 and 32 for details).  Figure 4: Principal component analysis. Genetic diversity and ancestry of the Scythian groups Haplogroups found in the Iron Age nomads are predominant in modern populations in both west (HV, N1, J, T, U, K, W, I, X) and east Eurasia (A, C, D, F, G, M, Y, Z). The mitochondrial haplotype diversity in our sample set ranges from 0.958±0.036 in the Tagar/Tes sample (group #7 in Fig. 2) up to 1.000±0.039 in the Early Sarmatians (#3; Supplementary Table 7). Using nuclear SNP data, we performed a principal component analysis33 (PCA) of 777 present-day west Eurasians26,34,35 onto which we projected the eight newly reported Iron Age Scythian samples as well as 167 other ancient samples from Europe, the Caucasus and Siberia from the literature17,34,36 (Fig. 4). The two Early Sarmatian samples from the West (group #3 in Fig. 2) fall close to an Iron Age sample from the Samara district34 and are generally close to the Early Bronze Age Yamnaya samples from Samara34 and Kalmykia17 and the Middle Bronze Age Poltavka samples from Samara34. The eastern samples from Pazyryk (#6), Aldy Bel (#5) and Zevakino-Chilikta (#4) are part of a loose cluster with other samples from Central Asia17, including those from Okunevo, Late Bronze Age and Iron Age Russia, and Karasuk. These samples contrast with earlier samples from the Eurasian Steppe belonging to the Andronovo17, Sintashta17 and Srubnaya34 groups, which overlap Late Neolithic/Bronze Age individuals from mainland Europe34,35 and are shifted downwards in the PCA plot towards the early farmers of Europe and Anatolia34.  |
|
|
|
Post by Admin on Mar 7, 2017 20:14:48 GMT
Since the PCA of west Eurasia in Fig. 4 does not allow one to examine the ancient samples in relation to contemporary East Asian populations, we also carried out PCA of all 2,345 modern individuals in the Human Origins dataset35, onto which we also projected the ancient individuals (Fig. 5). It is evident from this PCA that ancestry of the Iron Age samples falls on a continuum between present-day west Eurasians and eastern non-Africans, which is in concordance with the mitochondrial haplogroup analyses. The eastern Scythians display nearly equal proportions of mtDNA lineages common in east and west Eurasia, whereas in the western Scythian groups, the frequency of lineages now common in east Eurasia is generally lower, even reaching zero in four samples of the initial Scythian phase of the eight to sixth century BCE (group #1 in Fig. 2), and reaches 18–26% during later periods (sixth to second century BCE; #2 and #3) (Supplementary Table 7).  Figure 5: Principal component analysis. We used f4-statistics of the form f4(Test, LBK; EHG, Mbuti) and f4(Test, LBK; Han, Mbuti), which are zero for those Test samples that form a clade with LBK and positive for populations that have EHG- or Han-related ancestry, respectively. We plotted the results against each other, which resulted in a V-shaped pattern with Yamnaya at the apex (Fig. 6). The Iron Age Scythians are arrayed along a cline from Yamnaya to Ami (a population of East Asian ancestry that experienced no admixture), consistent with having ancestry from populations genetically similar to these two groups. We also computed statistics of the form f3(Test; Yamnaya_Samara, Han) to check whether a Test population has intermediate allele frequencies between Yamnaya_Samara and Han, which are used as proxies for possible source populations. Intermediate allele frequencies can only occur if the test population is a mixture of populations related to these two sources37. These statistics are significantly negative for all Scythians, demonstrating that admixture occurred (Supplementary Fig. 13).  Figure 6: Visualization of f-statistics results. We carried out ADMIXTURE analysis38,39 of 2,345 present-day humans35 genotyped on the Human Origins array35,37 and 175 ancient individuals on a set of 296,340 SNPs intersecting with those in the Human Origins array. The results for the ancient individuals are displayed in Fig. 7 for K=15, which has the highest log likelihood value (the complete analysis can be found in Supplementary Fig. 14). All steppe populations have ancestry components that are maximized in European hunter-gatherers (blue) and Caucasus hunter-gatherers from Georgia36 (green). One subset of the steppe populations (including Srubnaya, Sintashta and Andronovo) also have early farmer ancestry (orange), while a different subset (including all Iron Age samples) also have ancestry from a component (light blue) that is maximized in the Nganasan (Samoyedic people from north Siberian), and is pervasive across diverse present-day people from Siberia and Central Asia. Additionally, the Iron Age samples reveal an ancestral component that is maximized in East Asian populations (yellow), a type of ancestry that occurs at trace levels—if at all—among earlier steppe inhabitants, consistent with the observations from PCA and f-statistics about this type of admixture.  Figure 7: ADMIXTURE results for ancient populations. We modelled steppe populations as mixtures of the Early Bronze Age Yamnaya and the LBK farmers from central Europe or East Asians (represented by the Han Chinese). We applied the method of qpWave/qpAdm used in Haak et al.26, which provides a statistical test for the number of streams of ancestry into a Test population and allows one to estimate mixture proportions. In our application, we use five outgroups: Ust_Ishim40, Kostenki1441, MA142, Papuan and Onge. First we calculated whether Test and the Yamnaya from Samara could be descended from a single stream of ancestry. In the next step we included LBK farmers testing whether Test, the Yamnaya from Samara and the LBK farmers from central Europe could be descended from two streams of ancestry, in which case Test could potentially be modelled as a mixture of the other two populations. Our results show that the Iron Age Scythians and the Yamnaya are not descended from a single stream of ancestry (Supplementary Table 23) and furthermore, cannot be modelled as mixtures of the Yamnaya and the LBK (Supplementary Table 24). We therefore considered an alternative model in which we treat them as a mix of Yamnaya and the Han (Supplementary Table 25). This model fits all of the Iron Age Scythian groups, consistent with these groups having ancestry related to East Asians not found in the other populations. Alternatively, the Iron Age Scythian groups can also be modelled as a mix of Yamnaya and the north Siberian Nganasan (Supplementary Note 2, Supplementary Table 26). |
|
|
|
Post by Admin on Mar 8, 2017 20:16:15 GMT
Descendants of the Iron Age Scythians 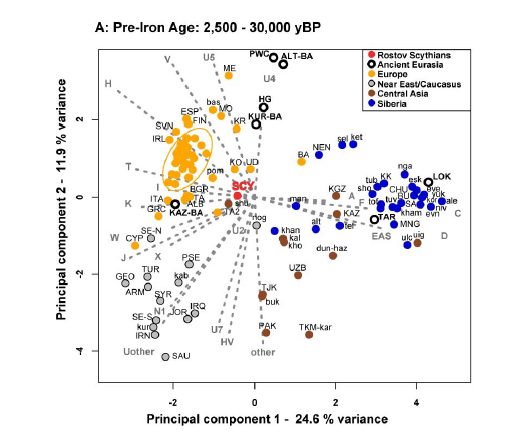 A multidimensional scaling (MDS) plot based on Reynolds’ distances (Supplementary Fig. 1) suggests that the ancient Scythian populations from the eastern and western part of the Eurasian Steppe are genetically closer to each other than are the modern populations of the respective regions. AMOVA analyses carried out for modern and ancient groups of the eastern and western steppe provided further support for this finding. We found FCT values to be higher between modern populations of the East and the West (FCT=0.0835) than between ancient populations of similar regions (FCT=0.0262). A continuity test was performed between the two Iron Age groups (‘West’ and ‘East’) and a large set of contemporary Eurasian populations (n=86, Supplementary Table 19). For western Scythian-era samples, contemporary populations with high statistical support for a genealogical link are located mainly in close geographical proximity, whereas contemporary groups with high statistical support for descent from eastern Scythians are distributed over a wider geographical range. Contemporary populations linked to western Iron Age steppe people can be found among diverse ethnic groups in the Caucasus, Russia and Central Asia (spread across many Iranian and other Indo-European speaking groups), whereas populations with genetic similarities to eastern Scythian groups are found almost exclusively among Turkic language speakers (Supplementary Figs 10 and 11).  Phenotypic markers Derived alleles of pigmentation markers that are under selection in Europeans are present in eastern and western Scythians, including individuals who are homozygous for the derived alleles at selected SNPs in the HERC2, SLC24A5 or SLC45A2 (ref. 43, 44, 45). At the two LCT loci associated with lactase persistence, the derived allele is observed only in heterozygotes, only in the eastern Scythian samples, and at low frequency (2–3%). The ancestral alleles at ADH1B rs3811801 and rs1229984 are nearly fixed in the Scythian dataset, as they are in modern Europeans (the derived alleles, which confer some resistance to alcoholism, are under selection in East Asians46,47). We observe the derived allele at rs3827760 in the EDAR gene in a single Pazyryk individual (#6 in Fig. 2). This EDAR derived allele, which is related to tooth and hair morphology, is selected and at high frequency in modern East Asians (87%)48, and very rare in modern Europeans (∼1%)48, although it has been observed in prehistoric hunter-gatherers from Sweden (7.9–7.5 kya)34. Thus, the results of the examination of phenotypic SNPs that show frequency differences between Europe and East Asia are consistent with gene flow across the steppe territory.  Our genomic analyses reveal that western and eastern steppe inhabitants possess east Eurasian ancestry to varying degrees. In our ADMIXTURE analyses we find an East Asian ancestry component at K=15 in all Iron Age samples that has not been detected in preceding Bronze Age populations in either western or eastern parts of the Eurasian Steppe. Another ancestral component that is maximized in the north Siberian Nganasan population becomes visible from the 2nd millennium BCE onwards in the eastern steppe (Okunevo, Karasuk, Mezhovskaya). This component appears later in all Iron Age populations but with significantly higher levels in the eastern steppe zone than in the West. These findings are consistent with the appearance of east Eurasian mitochondrial lineages in the western Scythians during the Iron Age, and imply gene-flow or migration over the Eurasian Steppe belt carrying East Asian/North Siberian ancestry from the East to the West as far as the Don-Volga region in southern Russia. In general, gene-flow between eastern and western Eurasia seems to have been more intense during the Iron Age than in modern times, which is congruent with the view of the Iron Age populations of the Eurasian Steppe being highly mobile semi-nomadic horse-riding groups.  In the East, we find a balanced mixture of mitochondrial lineages found today predominantly in west Eurasians, including a significant proportion of prehistoric hunter-gatherer lineages, and lineages that are at high frequency in modern Central and East Asians already in the earliest Iron Age individuals dating to the ninth to seventh century BCE and an even earlier mtDNA sample from Bronze Age Mongolia49. Typical west Eurasian mtDNA lineages are also present in the Tarim Basin16 and Kazakhstan8 and were even predominant in the Krasnoyarsk area during the 2nd millennium BCE31. This pattern points to an admixture process between west and east Eurasian populations that began in earlier periods, certainly before the 1st millennium BCE13,50, a finding consistent with a recent study suggesting the carriers of the Yamnaya culture are genetically indistinguishable from the Afanasievo culture peoples of the Altai-Sayan region. This further implies that carriers of the Yamnaya culture migrated not only into Europe26 but also eastward, carrying west Eurasian genes—and potentially also Indo-European languages—to this region17. All of these observations provide evidence that the prevalent genetic pattern does not simply follow an isolation-by-distance model but involves significant gene flow over large distances. Nature Communications 8, Article number: 14615 (2017) |
|
|
|
Post by Admin on Mar 12, 2017 20:08:35 GMT
 The enigmatic Scythians ruled the vast grasslands of Eurasia for a millennium. Legendary for their savagery on the battlefield, these horse warriors struck fear into their contemporaries in Greece, and in the Persian Empire. Unlike any of their neighbors in 700-something BC, or since, these unusual people had little use for the worldly trappings most identify with advanced civilizations. The Scythians left no major cities, but only the scattered grave mounds known as kurgans, which speckle the landscape of time from Mongolia to the Black Sea and Crimea. It is fair to say, no one yet knows from whence they came.  Whoever these fierce nomads were, the significance of the most recent archeological finds is amplified by the fact that the culture of the Scythians emerged almost exactly as the Bronze Age gave way to the Iron Age after the 1st Millennium BC. Assuming coincidences occur in flurries across recorded time, we might simply write off the people the Greeks feared as "barbarians" as only noteworthy barbarians that passed in time like so many others. An initial read of Herodotus' accounts today leads us to such a conclusion, but delving deeper we find a fabulously rich society, and a uniquely Russian one in many ways. The unusual ideology these Scythians possessed, the concept of "the people" or "true freedom", it was legendary in Persia and other bordering nations. Of the land, these Scythians had great regard. However, the tribes professed no real ownership of it, but rather domain over it. The distinction is subtle perhaps, but I believe the attitudes of these people correlate directly with some ideals held by current day Russians.  Herodotus, the father of history, said the Scythian women were in fact the original Amazons. Herodotus also recollected bizarre drug induced blood rituals, frenzied cannabis and opium fests where sacrifices may have been carried out. The Crimea finds in particular, tells us much of what the ancient Greek historian related, was more truth than fable. Taken in total, the fact the history of the Scythians is as remarkable as it is, and that it has been so overlooked, this creates a whole new series of questions. Who were these people really? How could history be revised so as to diffuse their impact, and why? We are only now beginning to uncover this impact, and the parallels with today's societies.  According to the historian, Scythians were born out of the Greek hero mating with a serpent/human goddess, one of the sons of this union becoming the first chieftain of this new nation. Legends of Scythians women fighting like the "Amazons" alongside their men, the rumors of being the lost tribesmen of Israel, there's no end to the mystique attached to this magnificent steppe culture. Why there are even studies that suggest the Scythians are the ancestors of the American plains Indians, the sole reason the Great Wall of China was built, the original people of Briton and Camelot even.  |
|

