|
|
Post by Admin on Mar 24, 2017 20:18:38 GMT
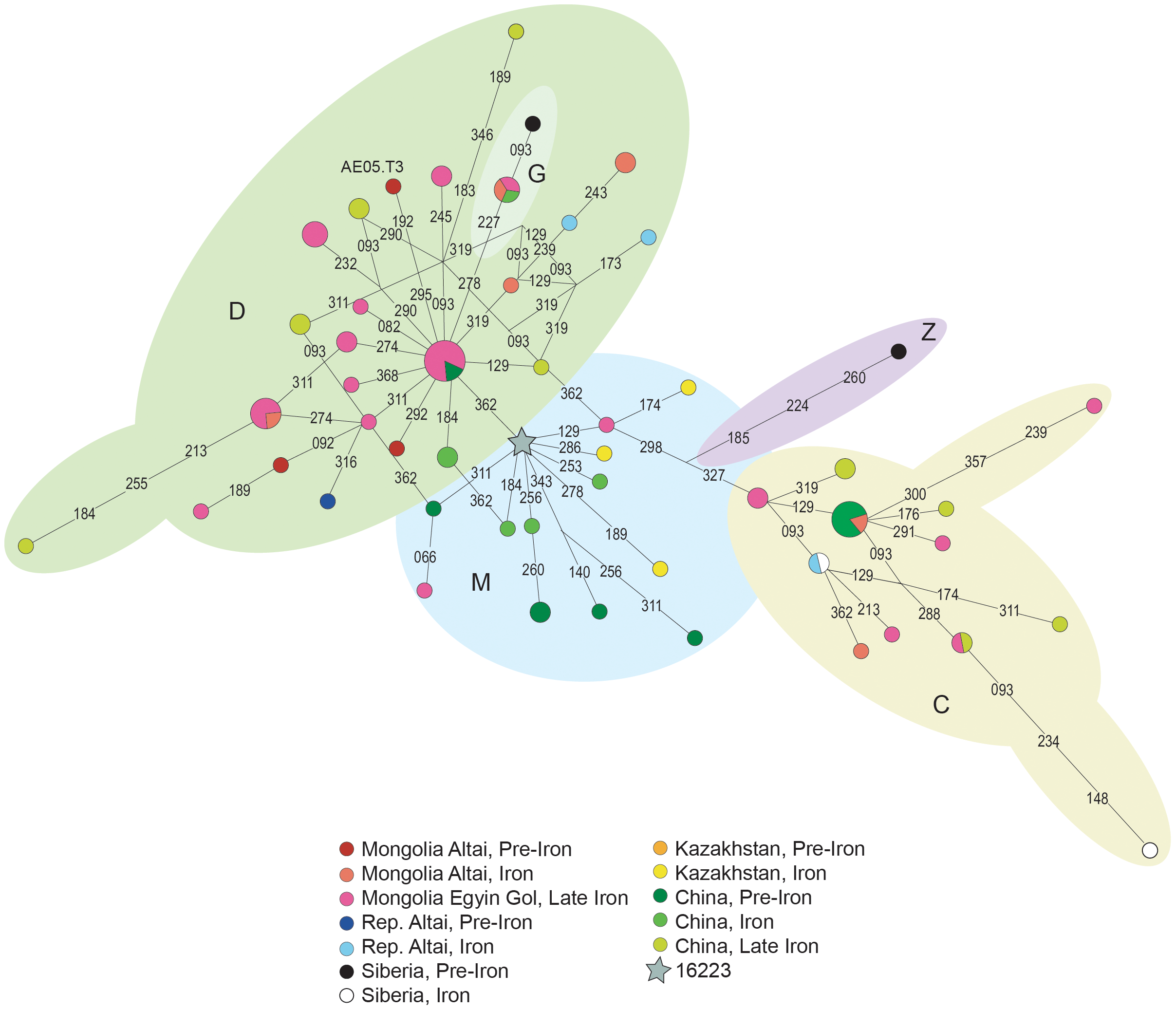 Figure 4. Median Joining Network of ancient M* haplogroup sequences. MtDNA sequences between positions 16051 and 16400, from ancient populations from the Mongolia (present study and [1], [2]), Russia [3], [4], [5], [6], Kazakhstan [7] and China [8], [9], [10] were used. Additional information concerning each population can be found in Table S2. Sequence raw data was analysed with Sequence Scanner v1.0 (Applied BioSystems) program and sequences were subsequently aligned with BioEdit software version 7.0.0 [30] in relation to the revised Cambridge Reference Sequence [31]. Samples were assigned to haplogroups using the combined information of HVRI and coding region variation following the phylogenetic classification updated by [32]. Haplogroups were clustered according to their geographic origin following [33], [34], [35]: West Eurasian haplogroups: R0: R0a’b, HV; N1; JT; UK; W and X. East Eurasian haplogroups: M: C, D, G, Z, M9, M10, M11, M13; A; B; F and N9a. South Asian haplogroups: M*, U2a-c, U9, R*, R1–R2, R5–R6, N1d. For comparative purposes, mtDNA sequences between positions 16051 and 16400, from ancient [10], [11], [18], [19], [20], [36], [37], [38], [39], [40] and modern populations [3], [41], [42], [43], [44], [45], [46], [47], [48], [49], [50], [51], [52], [53], [54], [55], [56], [57] of Eastern and Central Asia, were collected (See Table S2, for population codes and details about period, sample size and references). 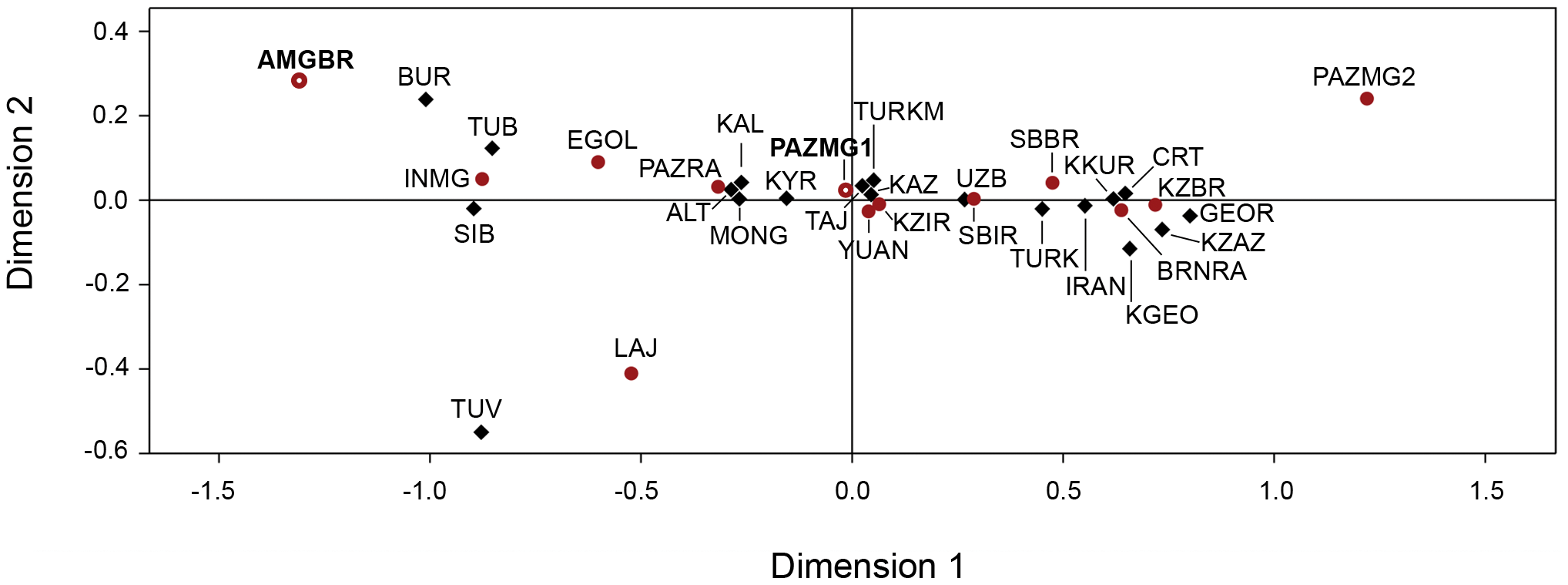 Figure 5. Multidimensional scaling representation of the Slatkin’s linearized FST pairwise genetic distance matrices between populations. Genetic distance based on HVRI variation of ancient and current Eurasian populations. Ancient populations (in red): AMGBR- Mongolia Altai Bronze Age, present study; PAZMG1- Mongolia Altai, Pazyryk, present study; PAZMG2- Mongolia Altai, Pazyryk; EGOL - Mongolia, Egyin Gol; PAZRA- Rep. Altai, Pazyryk; BRNRA- Rep. Altai, Neolithic and Bronze Age; SBBR- Siberia, Bronze Age; SBIR- Siberia, Iron Age; KZBR- Kazakhstan, Bronze Age; KZIR- Kazakhstan, Iron Age; LAJ- Lajia; YUAN- Xinjiang; INMG- Inner Mongolia. Current populations (in black): CRT- Crimean Tartars; TURK- Turks; KZAZ- Kurds Zazaki; KKUR- Kurds Kurmanji; IRAN- Iraqis; KGEO- Georgians Kurds; GEOR- Georgians; KYR- Kirgiz; UZB- Uzbeks; KAZ- Kazaks; TURKM- Turkmens; TAJ- Tajiks; MONG- Mongols; TUV- Tuvans; TUB- Tubalars; ALT- Altaians; BUR- Buriats; KAL- Kalmiks; SIB- Siberians. Additional information concerning each population can be found in Table S2. The majority of the retrieved sequences (58%) fit into East Eurasian lineages; namely to haplogroups A, C, D and G. On the other hand, 42% of the individuals belong to West Eurasian mtDNA haplogroups (J, K, HV, U, and T haplogroups) (Table 2). Considering the chronology of burials, the three Bronze Age samples represent three different haplotypes all of which can be classified as haplogroup D. For Iron Age samples, the same proportion (50%) of East and West Eurasian lineages were found (Table 2). Evidence of a perfect admixture between East Eurasian and West Eurasian lineages is also observed in other Iron Age populations from central Asia (Table 3 and Fig. 2B). On the other hand, in Neolithic and Bronze Age populations around 100% of the mtDNA lineages belong to East Eurasian or to Western Eurasian haplogroups, depending on the geographic location (Table 3 and Fig. 2A). The number of mtDNA haplotypes, Nei gene diversity [59] and nucleotide diversity [58], [59] (based on HVRI sequences) for the studied populations and for populations selected for comparison are presented in table 4. Our Bronze Age samples of Mongolian Altai (AMGBR) display the highest value of gene diversity. However, this value must be interpreted with caution since only 3 individuals were analysed and the error associated to this estimation is the highest reported. Concerning our Iron Age population (PAZMG1), the value of diversity is in the range of values observed in other ancient and present day populations from the same geographical area. Regarding the nucleotidic diversity, Bronze Age samples show the lowest values whereas Iron Age samples show high values. |
|
|
|
Post by Admin on Mar 25, 2017 19:51:32 GMT
 It is widely accepted that animal domestication (starting ∼5,000 YBP), particularly of the horse, gave the inhabitants of the Central Asian steppes the opportunity to expand geographically in different directions [12]. Historical records and archaeological data indicate that the early populations’ movements across the Eurasian steppe involved Indo-European-speaking people, being most probably in the Altai Mountains the eastern boundary. By the time of the 3rd century BC, Turkic-speaking peoples from the Altai region began to migrate westwards, replacing Indo-European languages in parts of Central Asia [12], [13], [14], [70]. Today, the Altai region is home to numerous Turkic-speaking ethnic groups, with a mixed mtDNA gene pool between West and East Eurasia [71]. However, the origin of this genetic admixture was prior to westward Turkic migrations and may be traced back to the Iron Age in Central Asia, or even earlier in South Siberia [40], [72]. With the present study, we have aimed to unravel the early contacts between European and Asian people in the Altai region by analyzing ancient mtDNA in human remains from Bronze and Iron Age burials of Mongolian Altai.  The high rate of success of the genetic analyses performed in the present study, evidenced by the amplification of both nuclear and mitochondrial DNA, suggests that the DNA of the samples is well preserved. In accordance with other authors [18], this is the expected result in samples that originated in cold and arid environmental conditions such as those found at most of the Altai sites. The agreement between morphological and genetic sex diagnoses, the concordant results obtained using coding and non-coding regions of mtDNA, the high haplotype diversity in sequence motifs, the concordant results between different extracts of the same sample, the clone results and the independent replication of some samples in another ancient DNA laboratory, all guarantee the authenticity of the obtained results [27], [28], [29]. Moreover, in terms of phylogenetic consistency, the observation of 16 different and phylogenetically plausible haplotypes among 19 ancient individuals and the coherence of the phylogenetic networks build further points towards the authenticity of the results [73].  Concerning Bronze Age samples from the Mongolian Altai mountains analyzed in the present study, 100% of the mtDNA lineages (3 different lineages from 2 archaeological sites) belong to East Eurasian haplogroups, an opposite profile to that detected in the West side of the Altai [18], [20]. On the other hand, in the Iron Age samples of Mongolian Altai, the same proportion (50%) of East and Western Eurasian lineages were found, evidencing a perfect admixture between East and Western Eurasian lineages as in other Iron Age populations from central Asia and Siberia [10], [18], [19], [20], [40]. Combined with the previous studies performed so far in the Altai region, our results suggest that the Altai represented a boundary to gene flow up to the beginning of the Iron Age and that during the Scythian period of the Altai (5th to 3rd century BC) there were demographic events in the region that led to a population admixture in both sides of the Altai.  Half of the shared haplotypes between ancient populations from Central Asia and South Siberia represent lineages present in both pre-Iron Age and Iron Age populations and all of these lineages have a west Eurasian origin. Moreover, considering both shared and non-shared haplotypes, it seems that the number of West Eurasian lineages does not increase in the Iron Age. These results allow us to hypothesise that the substrate of mtDNA lineages is already present in pre-Iron Age populations of the central Asia and that in the Iron Age (Scythian period) a population expansion lead to the admixture of pre-existing lineages. Thus, the admixture profile observed in the region during the Iron Age would not derive from a migratory movement from west to east, as has been hypothesised, but would represent a local population expansion in different directions. This population expansion, however, would be probably be a consequence of the introduction of new technology by the adoption of a new culture, supporting the idea of cultural transmission against the demic diffusion during Scythian period.  The Pazyryk groups analysed so far appear to be genetically homogeneous and they did not present significant genetic differences to current Altaians. These results suggest that roots of the current genetic diversity and admixture of the Altai region in Central Asia could be traced back to the Iron Age. González-Ruiz M, Santos C, Jordana X, Simón M, Lalueza-Fox C, Gigli E, et al. (2012) Tracing the Origin of the East-West Population Admixture in the Altai Region (Central Asia). |
|
|
|
Post by Admin on May 13, 2017 19:20:11 GMT
 Nomad Scythian herders roamed vast areas spanning the Central Asian steppes during the Iron Age, approximately from the 9th to the 1st century BCE (Before Common Era). These livestock pastoralists, who lived on wagons covered by tents, left their mark in the history of warfare for their exceptional equestrian skills. They were among the first to master mounted riding and to make use of composite bows while riding. A new study published in Science led by Professor Ludovic Orlando and involving 33 international researchers from 16 universities, now reveals the suite of traits that Scythian breeders selected to engineer the type of horse that best fit their purpose. The study took advantage of exceptionally preserved horse remains in royal Scythian burials, such as the site of Arzhan, Tuva Republic, where over 200 horses have been excavated but also at Berel', Kazakhstan, where no less than 13 horses were preserved in a single, permafrozen funerary chamber. Applying the latest methods in ancient DNA research, the researchers could sequence the genome of 13 Scythian stallions. These were 2,300-2,700 years old and included 11 specimens from Berel' and two from Arzhan. The researchers also sequenced the genome of one 4,100 year-old mare from Chelyabinsk, Russia, belonging to the earlier Sintashta culture, which developed the first two-wheeled chariots drawn by horses.  The DNA variation observed at key genes revealed a large diversity of coat coloration patterns within Scythian horses, including bay, black, chestnut, cream and spotted animals. Scythian horses did not carry the mutation responsible for alternate gaits, and as a consequence, were not natural amblers. However, some but not all individuals carried variants associated with short-distance sprint performance in present-day racing horses. This indicates that Scythian breeders valued animals showing diverse endurance and speed potential. "With the exception of two horses, none of the animals were related. It fits with Herodotus' depiction of Scythian funerary rituals, whereby sacrificed horses represented gifts from allied tribes spread across the steppes," says Dr. Pablo Librado, post-doctoral researcher at the Centre for GeoGenetics, University of Copenhagen, Denmark, and co-leading author of the study.  Importantly, none of the ancient horses analyzed in the study were inbred, which suggests that Scythian breeders succeeded in maintaining natural herd structures and did not perform selection through a limited number of valuable lineages. This contrasts with modern management practice where single stallions can be used to father hundreds of offspring. Patterns of genetic variation along the genome also revealed a total of 121 genes selected by Scythian breeders, most of which are involved in the development of forelimbs. This is reminiscent of the morphological indices measured on bones, and indicates that Scythian breeders selected horses showing more robust morphologies. "In this study we wanted to go beyond the myth of Scythians being aggressive warriors, drinking the blood of their enemies in skull mugs. We wanted to reveal the many facets of the exceptional relationship that these people developed with their horses," says Ludovic Orlando, Professor of Molecular Archaeology at the Centre for GeoGenetics, University of Copenhagen and CNRS Research Director at the AMIS laboratory, University of Toulouse. |
|
|
|
Post by Admin on May 14, 2017 19:21:55 GMT
 The genome data set generated in the study also reveals important lessons for the history of horse management, which started some 5,500 years ago, and animal domestication as a whole. By contrasting patterns of genetic variation in ancient and present-day horses, the authors found support for a significant demographic collapse during the last 2,300 years, which resulted in an important reduction of genetic diversity within horse domesticates. During the same time period, reproductive management has involved an increasingly reduced number of stallions, up to the point that, today, almost all domesticates virtually carry the same, or highly similar, Y-chromosome haplotype(s). "Many Y-chromosome haplotypes co-existed within Scythian horse populations. The first three millennia of horse domestication thus preserved a large diversity of male lineages. It only vanished within the last 2,000 years," adds Dr. Cristina Gamba, post-doctoral researcher at the Centre for GeoGenetics at the time of the study, and co-leading author of the study.  The authors also found that the demographic collapse and loss of Y-chromosomal diversity observed within the last 2,300 years was mirrored by a significant accumulation of deleterious mutations in the genome of the horse. As these mutations reduce the fitness of their carriers, it shows that the last two millennia of horse management have negatively impacted the horse. However, early domestication stages, as represented by the Sintashta and Scythian genomes, did not have such an impact. This contrasts with the Cost-of-domestication hypothesis, which posits a negative impact starting from early domestication stages. In the case of horse domestication, it is likely that the demographic collapse within the last 2,000 years reduced the efficacy of negative selection to purge out deleterious mutations, which could then accumulate in the horse genome. The domestication of the horse was a seminal event in human cultural evolution. Librado et al. obtained genome sequences from 14 horses from the Bronze and Iron Ages, about 2000 to 4000 years ago, soon after domestication. They identified variants determining coat color and genes selected during the domestication process. They could also see evidence of admixture with archaic horses and the demography of the domestication process, which included the accumulation of deleterious variants. The horse appears to have undergone a different type of domestication process than animals that were domesticated simply for food. Abstract The genomic changes underlying both early and late stages of horse domestication remain largely unknown. We examined the genomes of 14 early domestic horses from the Bronze and Iron Ages, dating to between ~4.1 and 2.3 thousand years before present. We find early domestication selection patterns supporting the neural crest hypothesis, which provides a unified developmental origin for common domestic traits. Within the past 2.3 thousand years, horses lost genetic diversity and archaic DNA tracts introgressed from a now-extinct lineage. They accumulated deleterious mutations later than expected under the cost-of-domestication hypothesis, probably because of breeding from limited numbers of stallions. We also reveal that Iron Age Scythian steppe nomads implemented breeding strategies involving no detectable inbreeding and selection for coat-color variation and robust forelimbs. Science 28 Apr 2017: Vol. 356, Issue 6336, pp. 442-445 DOI: 10.1126/science.aam5298 |
|
|
|
Post by Admin on Jul 9, 2018 18:41:54 GMT
 The Eurasian Steppe is a vast grassland region that stretches from the Carpathian foothills to Outer Mongolia. For millennia, the steppe was home to human populations that had a significant and long-lasting impact on the cultural history of the Eurasian continent. One such group that emerged at the beginning of the Iron Age had developed into an ethno-cultural agglomerate commonly referred to as the Scythians. The Scythians are best known from ancient Persian, Greek and Assyrian literary sources mainly for their nomadic warrior lifestyle, but they are also known to have actively practiced farming, pastoralism and may have been among the earliest peoples to master the art of horseback riding1. The territory occupied by contemporaneous nomadic and semi-nomadic Scythian groups with shared elements of material culture extended between the lower stretches of the Danube River in the west and the Yenisei River in the east2,3 (Fig. 1). Most archaeology and history researchers suggest that the core territory of the tribes designated as “Scythians” in historic literature of Antiquity covered the steppe and forest-steppe regions of the northern Black Sea (the North Pontic region, NPR) between the lower Danube and Don rivers4,5. Together with other contemporaneous groups they formed the ‘Scythian horizon’. These groups are collectively referred to by some researchers as Scytho-Siberians, who inhabited the steppe regions to the east of NPR Scythians and included Saka from Kazakhstan (7th–3rd century BCE)6,7 and Pazyryks (5th–3rd century BCE) from the Sayano-Altai region of Siberia2,8. The forest-steppe zone of the core Scythian territory in the NPR was settled by populations with agro-pastoral economy, while the nomadic and semi-nomadic Scythian tribes occupied the steppe regions adjacent to the northern Black Sea. Archaeological studies of the Scythian and pre-Scythian period sites in the forest-steppe zone of the NPR pointed towards autochthonous origins of local agro-pastoral Scythian populations. Those groups which first emerged between the middle 7th–3rd century BCE, may have been formed on the foundation of pre-existing groups belonging to such cultures as Srubnaya (Timber Grave) and Thracian Hallstatt9. The origins of nomadic Scythians in the steppes of Central Asia10 were supported by recent archaeological findings of elements of developed Scythian material culture in a series of kurgans in the western Sayan Mountains in southern Siberia dated to 9th–7th century BCE11.  Figure 1: Map of the Scythian core region and territories of their possible influences in 7th–3rd century BCE. The relationships between the groups of the Scythian horizon are not fully understood. Some researchers regard the NPR Scythians and Scytho-Siberians as one society on account of similar animal motifs on the products of their material culture, while others consider them to be different populations each having a distinct origin and geographic specificity yet sharing cultural traditions12. Moreover, the ancestral relationships between the NPR Scythians and local predecessor populations in the Ponto-Caspian region are neither fully resolved. Most of the current knowledge about the genetic relations among the populations belonging to the Scythian cultural horizon is based on studies of non-coding mitochondrial DNA (mtDNA) fragments from Scythian remains from the lower Don and southern Ural regions in Russia13 as well as Pazyryks from Altai and Inner Mongolia7,14,15,16,17,18. To date, only one complete mt genome of a Scythian individual from Russia has been published19. Mitochondrial lineages in the studied populations consist of an overlapping mix of haplogroups of east and west Eurasian descent, which does not clarify their ultimate origins. Here we aim to identify the maternal origin of the NPR Scythians and their genetic affinities to other contemporary Scythian groups through the analyses of complete mtDNA genomes. The investigated human remains were excavated from kurgans, crypts and ground burials in the main area of Scythian distribution, including territories in the lower Dniester, lower and middle Dnieper and Crimea. |
|

