|
|
Post by Admin on Mar 13, 2017 20:07:36 GMT
 Figure 1: Map of the Scythian core region and territories of their possible influences in 7th–3rd century BCE. The Eurasian Steppe is a vast grassland region that stretches from the Carpathian foothills to Outer Mongolia. For millennia, the steppe was home to human populations that had a significant and long-lasting impact on the cultural history of the Eurasian continent. One such group that emerged at the beginning of the Iron Age had developed into an ethno-cultural agglomerate commonly referred to as the Scythians. The Scythians are best known from ancient Persian, Greek and Assyrian literary sources mainly for their nomadic warrior lifestyle, but they are also known to have actively practiced farming, pastoralism and may have been among the earliest peoples to master the art of horseback riding1. The territory occupied by contemporaneous nomadic and semi-nomadic Scythian groups with shared elements of material culture extended between the lower stretches of the Danube River in the west and the Yenisei River in the east2,3 (Fig. 1). Most archaeology and history researchers suggest that the core territory of the tribes designated as “Scythians” in historic literature of Antiquity covered the steppe and forest-steppe regions of the northern Black Sea (the North Pontic region, NPR) between the lower Danube and Don rivers4,5. Together with other contemporaneous groups they formed the ‘Scythian horizon’. These groups are collectively referred to by some researchers as Scytho-Siberians, who inhabited the steppe regions to the east of NPR Scythians and included Saka from Kazakhstan (7th–3rd century BCE)6,7 and Pazyryks (5th–3rd century BCE) from the Sayano-Altai region of Siberia2,8. The forest-steppe zone of the core Scythian territory in the NPR was settled by populations with agro-pastoral economy, while the nomadic and semi-nomadic Scythian tribes occupied the steppe regions adjacent to the northern Black Sea. Archaeological studies of the Scythian and pre-Scythian period sites in the forest-steppe zone of the NPR pointed towards autochthonous origins of local agro-pastoral Scythian populations. Those groups which first emerged between the middle 7th–3rd century BCE, may have been formed on the foundation of pre-existing groups belonging to such cultures as Srubnaya (Timber Grave) and Thracian Hallstatt9. The origins of nomadic Scythians in the steppes of Central Asia10 were supported by recent archaeological findings of elements of developed Scythian material culture in a series of kurgans in the western Sayan Mountains in southern Siberia dated to 9th–7th century BCE11.  The relationships between the groups of the Scythian horizon are not fully understood. Some researchers regard the NPR Scythians and Scytho-Siberians as one society on account of similar animal motifs on the products of their material culture, while others consider them to be different populations each having a distinct origin and geographic specificity yet sharing cultural traditions12. Moreover, the ancestral relationships between the NPR Scythians and local predecessor populations in the Ponto-Caspian region are neither fully resolved. Most of the current knowledge about the genetic relations among the populations belonging to the Scythian cultural horizon is based on studies of non-coding mitochondrial DNA (mtDNA) fragments from Scythian remains from the lower Don and southern Ural regions in Russia13 as well as Pazyryks from Altai and Inner Mongolia7,14,15,16,17,18. To date, only one complete mt genome of a Scythian individual from Russia has been published19. Mitochondrial lineages in the studied populations consist of an overlapping mix of haplogroups of east and west Eurasian descent, which does not clarify their ultimate origins. Here we aim to identify the maternal origin of the NPR Scythians and their genetic affinities to other contemporary Scythian groups through the analyses of complete mtDNA genomes. The investigated human remains were excavated from kurgans, crypts and ground burials in the main area of Scythian distribution, including territories in the lower Dniester, lower and middle Dnieper and Crimea.  SCY192 Glinoe, Moldova 4th-2nd BCE 15–20 yrs XX H8c SCY193 Glinoe, Moldova 4th-2nd BCE n.a. XY U5a2a1 SCY196 Glinoe, Moldova 4th-2nd BCE 10–11 yrs n.r. W3a SCY197 Glinoe, Moldova 4th-2nd BCE n.a. XY U5a1a1 SCY300 Glinoe, Moldova 4th-2nd BCE 35–50 yrs XX H5b SCY303 Glinoe, Moldova 4th-2nd BCE 6–7 yrs XX U5a1a2b SCY305 Glinoe, Moldova 4th-2nd BCE 50 + yrs n.r. U5a2b SCY308 Glinoe, Moldova 4th-2nd BCE 20–35 yrs n.r. F1d SCY311 Glinoe, Moldova 4th-2nd BCE 35–50 yrs XX T2b SCY332 Glinoe, Moldova 4th-2nd BCE 30–35 yrs XX M10a1a1a SCY334 Glinoe, Moldova 4th-2nd BCE 20–35 yrs n.r. H5b SCY001 Svetlovodsk, Ukraine 4th BCE n.a. n.r. U5a1b SCY002 Svetlovodsk, Ukraine 4th BCE n.a. n.r. J1c2 SCY006 Starosillya, Ukraine 7th BCE n.a. XX D4j2 SCY011 Nesterivka, Ukraine 4th BCE n.a. XX A SCY012 Vapnyarka, Ukraine 4th-3rd BCE n.a. n.r. U5b2a1a2 SCY009 Starosillya, Ukraine 7th BCE n.a. XY J2b1a6 SCY010 Starosillya, Ukraine 7th BCE n.a. XX N1b1a SCY005 Simferopol, Ukraine 3rd-2nd BCE n.a. XX H n.a. - not available. n.r. - no result. Table 1: Description of studied individuals. Mitochondrial lineages in the NPR Scythians analyzed in this study appear to consist of a mixture of west and east Eurasian haplogroups. West Eurasian lineages were represented by subdivisions of haplogroup U5 (U5a2a1, U5a1a1, U5a1a2b, U5a2b, U5a1b, U5b2a1a2, six individuals total, 31.6%), H (H and H5b, three individuals total, 15.8%), J (J1c2 and J2b1a6, two individuals, 10.5%), as well as haplogroups N1b1a, W3a and T2b (one individual each, 5.3% each specimen). East Eurasian mt lineages were represented by haplogroups A, D4j2, F1b, M10a1a1a, and H8c (represented by a single individual), in total, comprising 26.3% of our sample set. The results of the low-resolution mtDNA screening at GVSU were consistent with whole mt capture results from AMU (see Supplementary Table S1). Polymorphism patterns uncovered in the coding and HVRI regions in specimens SCY009 and SCY011 identified their lineages to belong to haplogroups J and A respectively. Specimens SCY006 and SCY010 were assigned to the M* and N* clades respectfully. |
|
|
|
Post by Admin on Mar 14, 2017 20:12:55 GMT
 Figure 2: PC analysis based on mitochondrial DNA haplogroup frequencies. To trace genetic affinities between Scythians from present day Moldova and Ukraine (SCU) and other European and Asian ancient populations, their mt haplogroup frequencies were visualized in the space of principle components. The PCA plot of the first two components accounted for 43.4% of the total variance (Fig. 2). The SCU group was located in direct proximity to the central European Neolithic Corded Ware culture (CWC). It also grouped near a number of Bronze Age populations from eastern and central Europe (Srubnaya (SRU), Yamnaya (YAM) and Unetice (UNC)), as well as from central Asia (Bronze Age Kazakhstan (BAK)). Finally, k-means clustering (k value = 5), grouped SCU within a cluster further encompassing Scythians from Russia (SCR) and Tagar culture from southern Siberia (TAG). The Pazyryks from Mongolia (SCM) and Altai (SCA) were separated from SCU and grouped within the k-mean cluster consisting of Central and East Asian populations.  Figure 3: MDS plot based on FST calculated from complete mitochondrial genomes. Slatkin’s linearized pairwise FST values calculated on complete mt genomes and visualized using MDS (Fig. 3) and heatmap (Fig. 4) indicate that the studied Scythian samples are closest to the Srubnaya (SRU) (FST = 0.00; p < 0.05) and Yamnaya (YAM) (FST = 0.006; p < 0.05) populations (see Supplementary Table S5), followed by Unetice (UNC) (FST = 0.008; p < 0.05) and Corded Ware Culture (CWC) (FST = 0.023; p < 0.05). The European Neolithic Linear Ware culture (LBK) (FST = 0.043; p > 0.05) and Near Eastern Neolithic populations (NEN) (FST = 0.062; p > 0.05) appeared most distant to the individuals in our dataset. Correspondingly, in the resulted FST based MDS plot (stress value of 0.012), our Scythian group positioned proximately to Srubnaya (SRU), Yamnaya (YAM) and Unetice (UNC) populations and distantly to Near Eastern and European Neolithic (NEN, LBK) and Near Eastern hunter-gatherer (HGNE) populations (Fig. 3).  Figure 4: Heatmap of FST and geographic distribution. |
|
|
|
Post by Admin on Mar 15, 2017 20:15:11 GMT
 Figure 5: Median network of U5 haplotypes. Considering the high number of Scythian individuals identified as belonging to haplogroup U5 (31.6%), we conducted median network analysis of published ancient haplotypes belonging to U5a and U5b subdivisions and ranging from the Mesolithic (excluding hunter-gatherers predating last glacial maximum) to the Iron Age (Fig. 5). Haplotypes used in the median network analysis are described in detail in Supplementary Table S4. The Scythians in the U5a cluster were located centrally in the median network, near the U5a ancestral node. The U5a Scythian samples (SCU) grouped together with Mesolithic hunter-gatherers from Sweden (haplotypes 3, 4 and 5), Germany (8), Russia (10) and France (45 and 46). Non-Mesolithic samples in those clusters were less numerous and included Srubnaya (SRU) (57) and Yamnaya (YAM) (13 and 14) from Samara region and one representative of Karasuk culture from the Altai territory (central-south Siberia) (24). A single U5b Scythian individual (31) was distant from the East European populations and was placed basally to a broader cluster composed of west hunter-gatherers (HGW) from France (41) and Germany (47) as well as chronologically younger samples from the Middle Neolithic (MNE) and Unetice (UNC) (both from Germany) (15 and 53, respectively). On the basis of published data concerning the phylogeography of mt lineages distribution in ancient populations of Europe and Asia, the 19 complete mt genomes of the NPR Iron Age Scythians produced in this study fall into three main groups of different ancestry. The first group of mt lineages is represented by U5 haplotypes that are considered to be a European Hunter-Gatherer genetic component44,45. The second group comprises haplotypes belonging to H, J, T, W and N1b, ultimately connected to the genetic package of the early Neolithic farmers44,46,47, and the third group includes A, D, M10 and F mt lineages considered to be of East Eurasian origin48,49,50,51. Representatives of the U5 haplogroup account for one third of the mt haplotypes identified in analyzed Scythians. These ultimately relate to West Eurasian hunter-gatherers, whose descendants extended throughout the European subcontinent and into East Eurasia from the Mesolithic to the Bronze Age. We have combined the known diversity of the prehistoric U5 lineage from different sources19,45,46,47,52,53,54,55,56 into a median network, which allowed for identification of candidate source populations contributing to the U5a diversity in NPR Scythians. Although the network approach is still limited by the small number of available samples, our results indicate that the source populations with the most closely related haplotypes are the Bronze Age Srubnaya (SRU) (1900-1200 BCE) and the earlier Yamnaya (YAM) (3300-2700 BCE) from the Ponto-Caspian region and Bronze Age populations from the Altai Mountains, such as Karasuk (1500-800 BCE). Diverse U5a-carrying populations of the steppe such as Yamnaya, Srubnaya and Scythians shared the nomadic lifestyle, with the economic foundations transforming from wild game exploitation in the Mesolithic to pastoralic animal husbandry in the Bronze Age. Furthermore, both Yamnaya and Srubnaya were part of the Kurgan culture phenomenon57 with the Scythian cultural horizon being the most representative for the kurgan building tradition. The second group of identified mt genomes in the NPR Scythians is comprised of lineages that suggest associations of the NPR Scythians with the Neolithic European farming groups. Although the mt lineage composition of analyzed Scythians significantly differs from that seen in NEN and LBK groups (FST = 0.06, FST = 0.04, respectively, p > 0.05), particular lineages such as J1c2, T2b, H/H5, H, W and N1b1a ultimately go back to the earliest European farmers. Lineages of J1c, H5 and T2b belong to the Neolithic farming package of mtDNA haplogroups which have been found in most Neolithic and Bronze Age European populations19,44,47,55,58. Noteworthy, individuals belonging to the N1b1a have been found in the Neolithic Anatolia19,59, but the lineage has not appeared in any other Eurasian Neolithic, Bronze or Iron Age populations to date. Therefore, the presence of N1b1a in Scythians could be attributed to population migrations along the southern boundaries of the Ponto-Caspian region60. 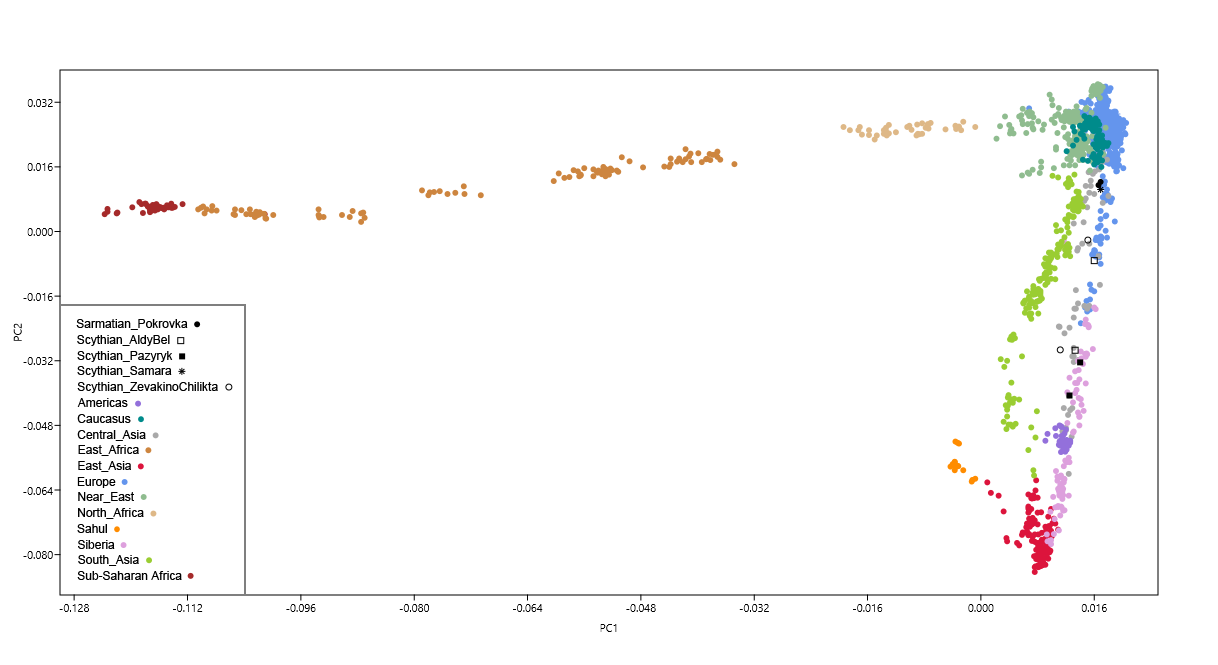 Mitochondrial lineages T2b, W3a and J2b1a6 have been identified in representatives of the Bronze Age groups such as Srubnaya (T2b, J2b1a, H5)19, Sintashta (J2b1a, J1c)55, Yamnaya (W3a and T2b)47,61 and Mezhovskaya (J2b1a)55 along with East European Eneolithic Trypillian culture (T2b)62, suggesting genetic continuity of these lineages from at least the Bronze Age or even Neolithic times in NPR region. This is further supported by published data19 which showed that almost one fifth of the genetic makeup of the Late Bronze Age Srubnaya people of the Ponto-Caspian region is of the Early European Farmer or Anatolian Neolithic ancestry possibly resulting from the admixture of populations related to Early European Farmers and Yamnaya. Thus, if the NPR Scythians are the descendants of populations related to Srubnaya, the origin of the identified farming lineages would likely be within the steppe/forest-steppe region between the NPR and southern Ural. According to previous genomic studies47,55 the CWC people are likely to have arisen from Yamnaya background. Thus, genetic affinities of the NPR Scythians to the Yamnaya people might also explain their close genetic similarity to the CWC reflected in PCA and FST results. Close genetic relations of the NPR Scythians and Srubnaya are supported by the FST analyses revealing no significant differences between these two populations (Figs 3 and 4, Supplementary Table S5). AMOVA further corroborated these results showing that the combination of Scythians and Srubnaya results in the lowest genetic variability among populations in a group and the highest variability between groups (Supplementary Table S6). Our results support the archaeological interpretations concerning the origin of the Scythians. One of the versions of this theory suggests that the Srubnaya people have migrated in several waves from the Volga-Ural region to the NPR during the second half of the second millennium BCE where their descendants gave rise to the Scythians around the 7th century BCE63. The third group of lineages identified in the NPR Scythians is derived from East Eurasian ancestry. Since, to our knowledge, there is no evidence of agricultural subsistence in East Eurasian Scythians, these lineages should be considered to be genetic components associated with nomadic populations. Mitochondrial haplogroups such as A, D and F have already been found in samples from the Mesolithic, Neolithic, Bronze and Iron Age southern Siberia and the Altai7,14,15,16,18,55,64,65,66,67. Notably, haplogroup M10 found among Scythians from Glinoe, is present not only in far East Asia but also in modern populations of the Altai68. Among ancient populations, the M10 lineages have been found in Chinese specimens from southern Xinjiang48 (8th -1st century BCE) and Xiongnu69 (3rd century BCE). Furthermore, haplogroup M have been identified in Pleistocene individuals from western Europe but it is thought that these lineages disappeared due to bottleneck effect in the Last Glacial Maximum56. Haplogroup H8c, identified in the NPR Scythians likely belongs to East Eurasian lineages as well. 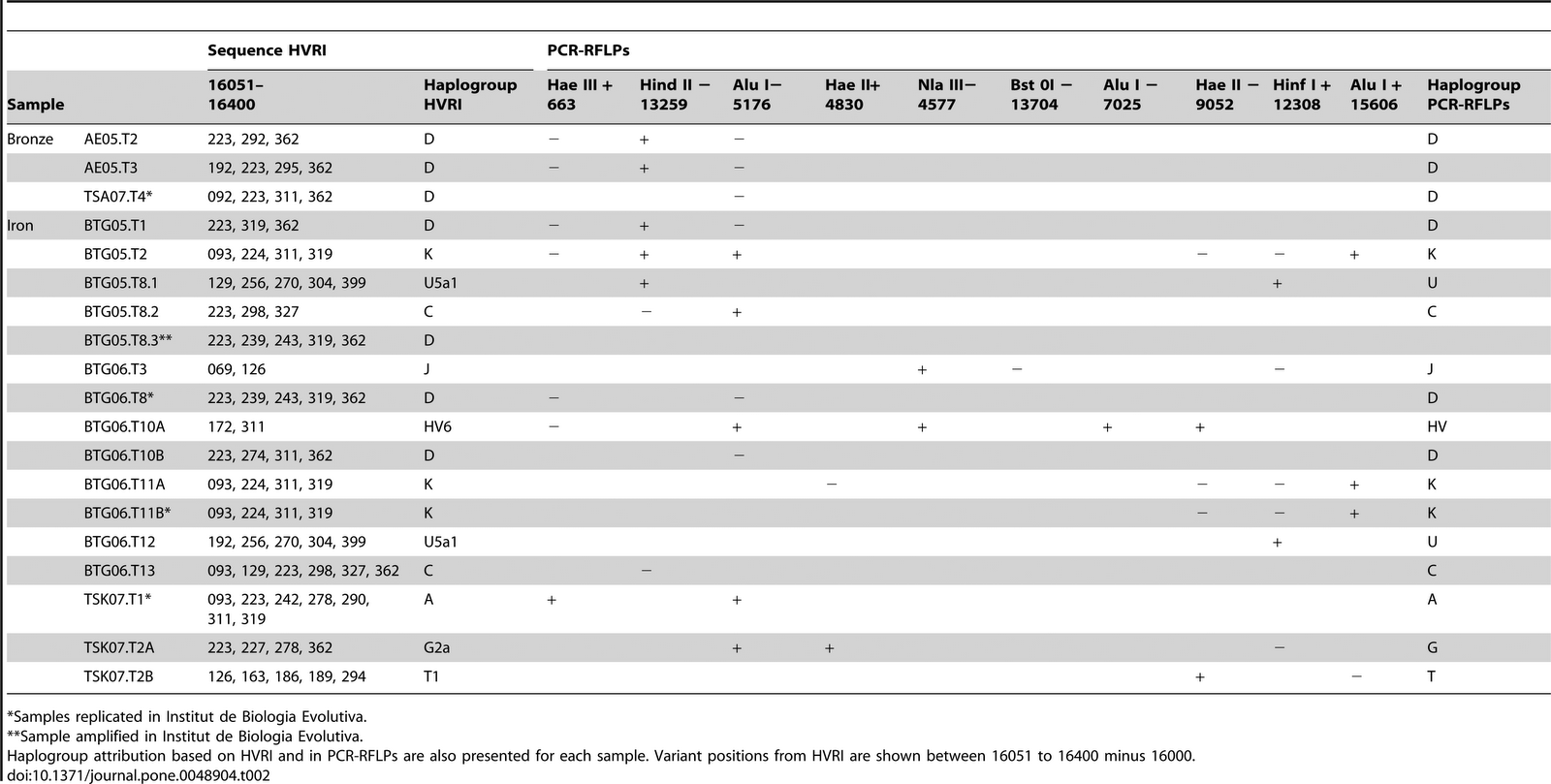 Sequence analysis of 830 modern Eurasian mt genomes suggested a distinct phylogeographc history for H8, with a clustering of Near Eastern and Central Asian haplotypes of H8 and a pronounced presence of carriers of H8c in the Altai region70. It was further hypothesized that the distribution of this lineage could have been facilitated by nomadic migrations along the NPR coast70. Although our PCA analysis showed Altai Pazyryk (SCA) to be distant from the NPR Scythians (SCU) (Fig. 2), it must be emphasized that the presence of haplogroups A and H8c in the analyzed population connects NPR Scythians to the Altai and identifies this region as a possible source of these East Eurasian mt lineages. The only Scythian mt genome from southern Urals published thus far also belonged to an East Eurasian lineage, G2a19. The Y chromosomal lineage (R1a1a1b2a2a) reconstructed for this individual was supposedly characteristic for members of the Srubnaya culture19 which additionally supports our conclusions concerning close genetic links between Scythians and people related to the late Bronze Age Srubnaya. Scythians from Rostov-on-Don as well as Pazyryks from Altai and Inner Mongolia were carriers of mixed east and west Eurasian lineages, with the dominant presence of the latter at 62.5% and 53.3%, respectively7,13,14,15,16,17,18. Mitochondrial haplogroup analyses of the NPR Scythians from this study and those from Rostov-on-Don and Pazyryks from Altai and Inner Mongolia, reveal that, for the most part, the same lineages are found in all three groups and are often singularly represented in each group. Juras, A. et al. Diverse origin of mitochondrial lineages in Iron Age Black Sea Scythians. Sci. Rep. 7, 43950; doi: 10.1038/srep43950 (2017). |
|
|
|
Post by Admin on Mar 22, 2017 20:12:18 GMT
 Figure 1. Pazyryk burials from the Altai Mountains. A. Geographical location of Pazyryk culture sites in the Altai regions of South Siberia, Kazakhstan and Western Mongolia. B. Pazyryk burial from Baga Turgen Gol site, Bayan-Ölgiy province, Western Mongolia. The Pazyryks have traditionally been associated with the Eastern Scythians. Scythians, whose history is well-known from the ancient texts of Herodotus (484–425 BC), was the name that the Greeks gave to a number of separate Indo-European-speaking nomadic groups living in the region encompassing the Pontic-Caspian steppe (in Eastern Europe) and the steppe of Central Asia. The Scythian culture (7th-2nd century BC) flourished in this region from local Indo-European peoples that emerged at the Pontic-Caspian steppe in about 2000BC and expanded eastward until they reached the Altai Mountains. Advances in technology that favoured mounted nomadic pastoralism were the triggers for the expansion of Scythian culture across the Western Eurasian steppe. The end of Scythian period might be related with the westward migrations of nomadic Turkomongolian tribes coming from East Asia since the 3rd century BC, which marked the end of Indo-European domination of the steppe [12], [13], [14], [15]. Archaeological findings, almost entirely provided by burial site discoveries, documented that the Scythians had European morphological features [7], [8], [12]. However, several recent works focusing on ancient mitochondrial DNA (mtDNA) of Eastern Scythian burials [9], [10], [11], [16], [17], [18], [19] revealed that this population has a mixed mtDNA composition of West and East Eurasian lineages. This is particularly interesting for the timing of the early contacts between European and Asian people in Altai because all ancient DNA samples analysed so far from Central Asia belonging to a period before the Iron Age bore West Eurasian lineages [18], [20].  These molecular data raise two likely hypotheses for the origin of the genetic diversity and admixture among the Iron Age inhabitants of the Altai: 1) people holding west Eurasian lineages arrived at Altai Mountains with the eastward migration of Scythians and, once settled, they began to establish relationships with the neighbouring communities from East Asia holding East Eurasian lineages; 2) this was the result of the admixture between the native people inhabiting either sides of the Altai Mountains (people with West Eurasian lineages in Western Altai and East Eurasian lineages in the Eastern Altai), as a result of a demographic expansion during the Scythian period. Hence, the second hypothesis would provide support to the cultural transmission against the demic diffusion during the Scythian period. The skeletal remains unearthed by our team from Bronze and Iron Age tumuli in the Mongolian Altai offer a unique opportunity to get further insights into the ancient Altaians. Here, we aim to shed light on the origin of the current east-west population admixture in Central Asia, specifically in the Altai region, by analyzing the mitochondrial DNA lineages of these skeletons. Owing to the clear geographical structuring of the East and West Eurasian lineages in Central Asia population, mtDNA is particularly suited for the study of admixture in this region [18]. This study may contribute to providing early evidence of population admixture between European and East Asian people, as well as the underlying causes behind this demographic event in the Eurasian steppes. |
|
|
|
Post by Admin on Mar 23, 2017 20:13:56 GMT
For each sample, the mtDNA hypervariable region I (HVRI) was amplified and sequenced, and coding region informative polymorphisms for haplogroup assignment were analysed by PCR-RFLPs. The PCR reactions were carried out in a final volume of 50 µl and Taq polymerase (Bioline®) was used. Each PCR reaction consisted of an initial denaturation step (5 min at 94°C) followed by 39 cycles of PCR (50 s at 94°C, 1 min at annealing temperature depending on the region to be amplified, and 1 min at 72°C) and a final extension step of 5 min at 72°C, or of 10 min if the amplified segment was to be cloned. Amplified fragments were then visualized with Ethidium Bromide staining in a 3% agarose gel [23]. To analyse the HVRI, four overlapping fragments were used (Table S1). These were subsequently sequenced and cloned. Sequence reactions were carried out using the sequencing kit BigDye Terminator v.3.1 (Applied Biosystems, Carslbad, USA) according to the manufacturer’s specifications and run in an ABI 3130XL sequencer.  Figure 2. Spatial frequency distribution maps of East Eurasian lineages. A- Pre-Iron Age period; B- Iron Age period. Frequency values and detailed information for populations 1–8 are shown in table 3. 1- Mongolia (Altai), 2- Gorny Altai, 3- West Kazakhstan, 4- Central Kazakhstan, 5- South Kazakhstan, 6- East Kazakhstan, 7- SW Siberia, 8- Mongolia (Egyin Gol). For all samples, the fragment containing the majority of HVRI mutations was cloned using the Topo TA Cloning® kit (Invitrogen, Carslbad, USA) following the manufacturer’s instructions. The colonies were harvested and subjected to PCR with M13 universal primers; for each sample 10 inserts of the right size were subsequently sequenced.  Table 3. Frequency of West and East Eurasian haplogroups in ancient Eurasian populations prior to Iron Age and from the Iron Age. Sex Determination For genetic sex determination, X and Y Amelogenin loci and the SRY gene (sex-determining region Y gene) were analyzed using primers and conditions described respectively by Beraud-Colomb et al. [24] and Santos et al. [25].  Figure 3. Median Joining Network of ancient N* haplogroup sequences. MtDNA sequences between positions 16051 and 16400, from ancient populations from the Mongolia (present study and [1], [2]), Russia [3], [4], [5], [6], Kazakhstan [7] and China [8], [9], [10] were used. Additional information concerning each population can be found in Table S2. |
|

